La circulation de matière au sein de la plante à fleurs
Vidéo en bas de page ^^
Les feuilles ne peuvent réaliser la photosynthèse sans l’eau prélevée par les racines. Les cellules de ces dernières ne pratiquent pas la photosynthèse et ont donc besoin des molécules produites par les feuilles. Il y a donc une nécessaire circulation de matière qui s’effectue dans la plante.
I Une circulation de sève brute des racines vers les feuilles.
Les racines sont le lieu de production de la sève brute (eau et sels minéraux) acheminée jusqu’aux feuilles pour fabriquer de la matière organique. Cette circulation peut être mise en évidence très simplement en faisant tremper des pétioles de céleri dans de l’eau contenant un colorant vital bleu. Au bout de quelques jours, les feuilles deviennent bleues. Une coupe du pétiole met en évidence des vaisseaux colorés en bleu. On retrouve ces vaisseaux dans la feuille et dans la tige. On les appelle les vaisseaux du Xylème, du grec du « xylon » qui veut dire « bois ».
Document 1 : Dispositif expérimental permettant de mettre en évidence une circulation ascendante de sève et résultats (feuille et coupe de pétiole)



Les vaisseaux du xylème transportant la sève brute des racines vers les feuilles, sont constitués de cellules mortes très allongées présentant des parois cellulosiques épaissies par des dépôts de lignine. Ces cellules sont empilées les unes sur les autres pour former un tube, elles sont donc dépourvues de parois transversales à leurs extrémités. L’association de ces cellules forme des vaisseaux véhiculant la sève brute. Ils sont colorables en vert par le colorant « carmino-vert de Mirande ». Ils permettent le soutien de la plante (lignification = formation de bois).
Ces épaississements de lignine peuvent prendre des formes différentes.
La lignine est une longue molécule qui peut s’enrouler formant une sorte de ressort rigidifiant le tube : on observe alors des vaisseaux spiralés. Il est possible d’observer également des vaisseaux réticulés et des vaisseaux ponctués selon la position que prend la lignine dans la paroi.
Document 2 : Vaisseaux du xylème du céleri mis en évidence par du bleu de méthylène.

II. Une circulation de sève élaborée des feuilles vers le reste de la plante.
Les feuilles sont le lieu de la production de matière organique et notamment des sucres. Le glucose produit est stocké sous forme d’amidon dans les chloroplastes et dans les amyloplastes car cette molécule est trop grosse pour circuler dans les vaisseaux. Pour permettre la distribution du sucre à l’ensemble de la plante, la cellule va associer un glucose et un fructose pour former un diose, le saccharose facilement transportable par la sève.
Document 3 : Structure de l’amidon
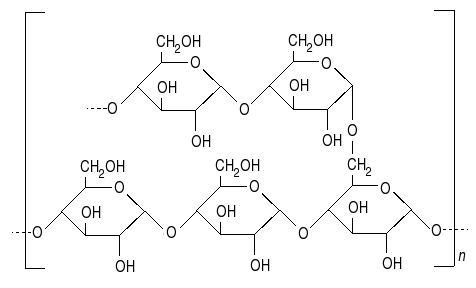
Source : Structure de l'amylopectine.JPG par Laranounette via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Structure_de_l%27amylopectine.JPG
Document 4 : Formation d’une molécule de saccharose
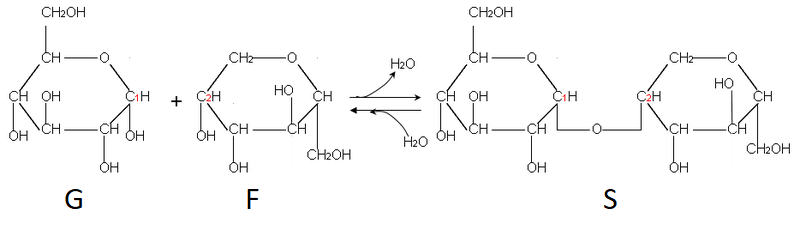
Source : Formation du saccharose.PNG, par Belgarath007 via Wikimédia Commons, CC-BY-SA-3.0-migrated, https://commons.wikimedia.org/wiki/File:Formation_du_saccharose.PNG?uselang=fr
Les feuilles peuvent également utiliser le glucose produit pour fabriquer d’autres molécules. Ainsi la sève formée au niveau des feuilles est riche en matière organique élaborée par les cellules chlorophylliennes, d’où le nom de « sève élaborée ». La matière organique (eau, sucres, acides aminés, ions minéraux…) est donc distribuée à l’ensemble de la plante via la sève élaborée.
Cette circulation de sève peut être mise en évidence par radioactivité. En effet, il suffit d’offrir à la plante du CO2 marqué au carbone 14 radioactif pour suivre le déplacement des sucres formés à partir de celui-ci. Au bout de 24h d’exposition au niveau foliaire, la radioactivité se retrouve dans les racines de la plante.
Document 5 : Vaisseaux du phloème et cribles

Les faisceaux du phloème, de petit diamètre transportent la sève élaborée des feuilles vers le reste de la plante. Ils sont constitués de nombreuses cellules vivantes accolées ensemble à leurs extrémités par une paroi cellulosique percée de petits trous : les cribles. Ces vaisseaux sont colorés en rose par le carmin-vert d’iode.
Document 6 : Vaisseaux du phloème et cribles
Ce système de vaisseaux à parois transversales perforées a pour effet de ralentir la descente de la sève, ce qui n’aurait pas été le cas avec une structure type xylème. Cela permet une distribution homogène à l’ensemble de la plante. Ce transport se fait donc de manière descendante mais aussi latérale. La vitesse de circulation de la sève élaborée est donc inférieure à celle de la sève brute. Cela ne pose pas de problème « d’embouteillage » dans les vaisseaux descendants car la majeure partie de l’eau de la sève brute ascendante est perdue par évaporation au niveau des feuilles : il y a donc peu d’eau de la sève brute qui repart dans le circuit descendant après passage dans la feuille.

III. Répartition des vaisseaux dans les organes
Les vaisseaux du phloème et du xylème ont des positions relatives (les uns par rapport aux autres) qui diffèrent selon les organes.
Dans la tige et dans la feuille, xylème et phloème sont regroupés, en position superposée, formant ce qu’on appelle un faisceau criblo-vasculaire. Dans la feuille, au niveau de la nervure centrale, le phloème est toujours situé du côté de l’épiderme inférieur alors que le xylème est situé du côté de l’épiderme supérieur.
Document 7 : Coupe de feuille de maïs et son faisceau criblo-vasculaire de la nervure centrale
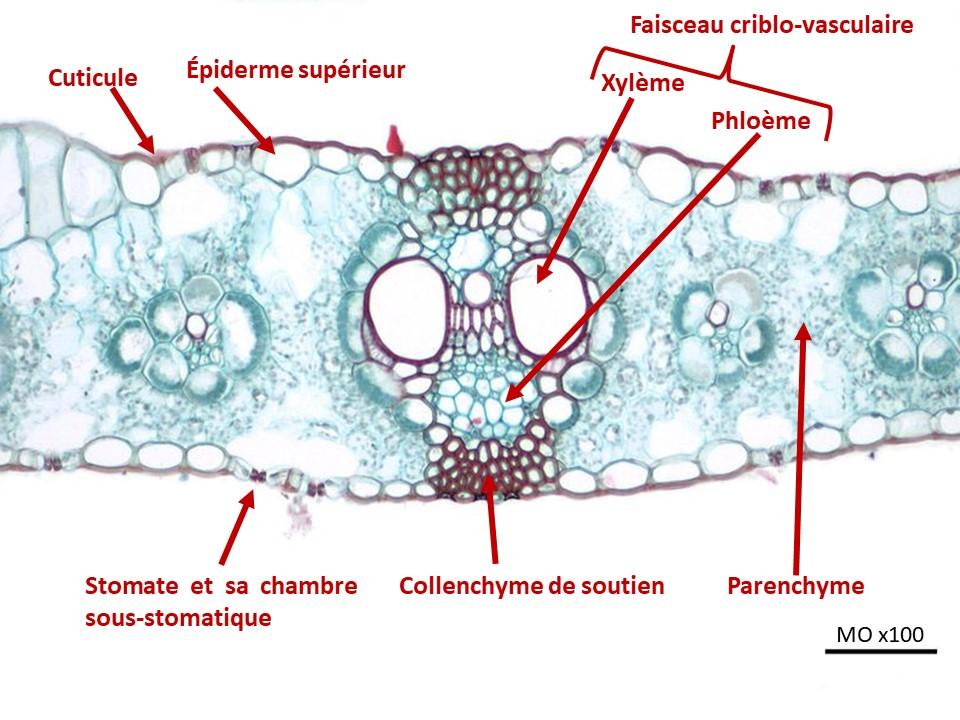
Source : Zea mays leaf.jpg, par Jon Houseman, via wikimedia commons, CC-BY-SA-4.0, modifié par Sandra Rivière , https://commons.wikimedia.org/wiki/File:Zea_mays_leaf.jpg
Du fait de sa position dans la feuille, arrivé dans la tige, le phloème passe en position externe alors que le xylème passe en position interne.
Document 8 : Disposition des vaisseaux dans une plante vue en coupe

Animation : http://viasvt.fr/circulation-seves/circulation-seve.html
Dans une racine, phloème et xylème sont regroupés dans un cylindre central délimité par une couche de cellules imperméable, l’endoderme. Dans ce cylindre central, les vaisseaux du xylème et du phloème sont en position alterne.
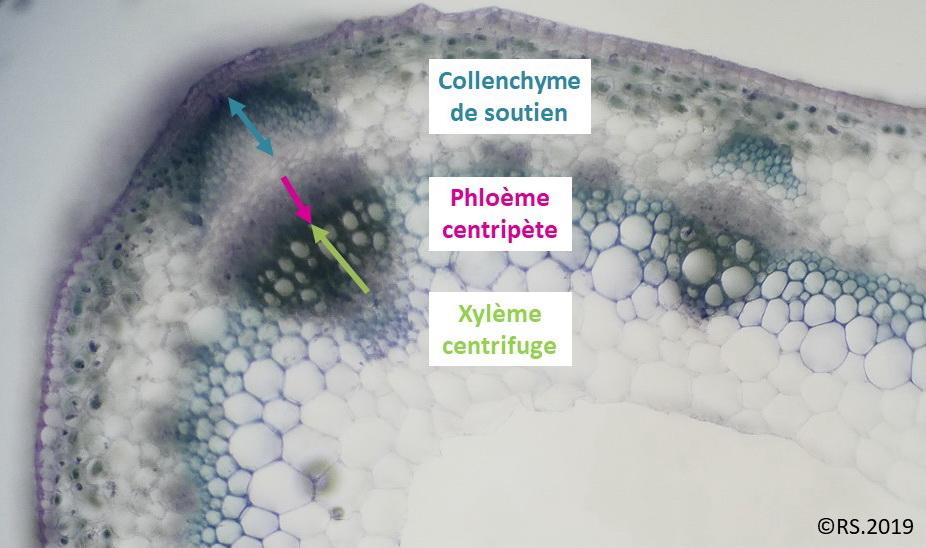
Que ce soit dans une racine ou une tige, la différenciation du phloème est toujours centripète : le phloème va se former de l’extérieur vers l’intérieur. Par contre le xylème a une différenciation centripète dans la racine (il se développe en direction du centre de l’organe) alors que dans une tige il a une différenciation centrifuge (il se développe en s’éloignant du centre de l’organe).
Document 9 : Cylindre central d’une racine : phloème et xylème sont alternes

Document 10 : Un des faisceaux conducteurs d’une tige de petit pois : phloème centripète et xylème centrifuge
IV. Circulation de l’eau et de la sève
Les solutions du sol traversent donc la paroi hydrophile des poils absorbants et remontent en direction des vaisseaux du cylindre central de la racine en raison d’une pression osmotique de plus en plus élevée (milieu de plus en plus hypertonique).
Plusieurs voies de circulation sont possibles :
- la voie du symplaste ou voie symplasmique : il existe un réseau de petits tunnels connectant les cytoplasmes des cellules et appelés « plasmodesmes ».
- La voie de l’apoplaste ou voie apoplasmique : on appelle apoplaste, l’ensemble de l’intérieur des cellules mortes, les parois cellulaires et les interstices entre les cellules, soit l’ensemble des espaces morts présents dans un tissu.
- La voie transcellulaire, c’est à dire à la fois par l’apoplasme et par le symplasme : l’eau et les sels minéraux traversent les membranes plasmiques et les parois cellulosiques.
Document 11 : Différentes voies de circulation de l’eau
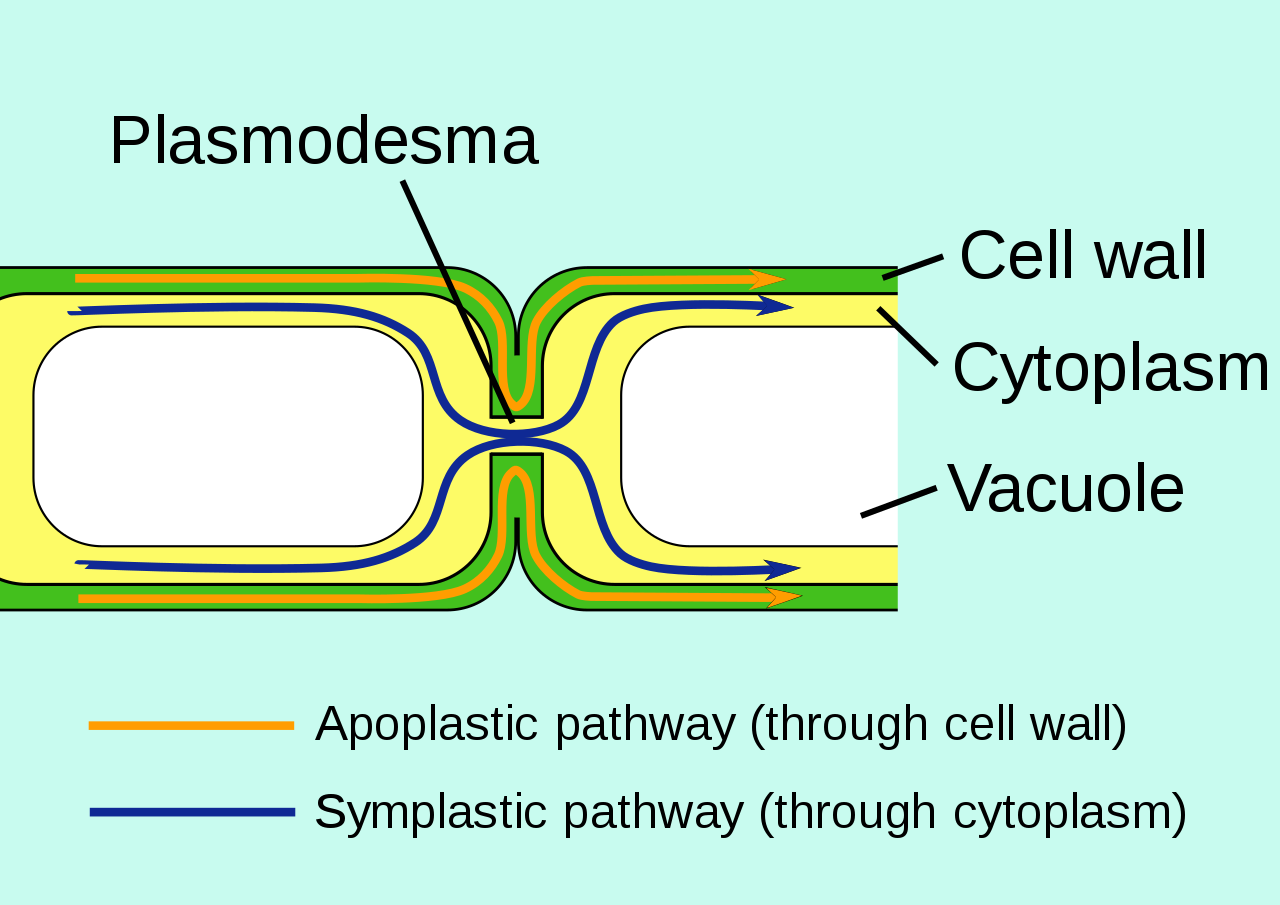
Source : Apoplast et symplast pathways.svg, par Jackacon , vectorisé par Smartse domaine publique, via Wikimedia commons, https://commons.wikimedia.org/wiki/File:Apoplast_and_symplast_pathways.svg
L’eau traverse la paroi cellulaire du poil absorbant au niveau de canaux protéiques appelés aquaporines mais ne rentre pas dans le cytoplasme de celui-ci. L’eau circule de cellule en cellule par la voie apoplasmique, c'est-à-dire entre les membranes et les parois cellulaires. Elle va d’abord circuler dans le parenchyme cortical (cortex) puis devoir traverser l’endoderme avant d’arriver aux vaisseaux.
Or l’endoderme est constitué de cellules parallélépipédiques avec des parois axiales minces et des parois radiales épaissies, constituées de cire (la subérine), le tout formant autour de la cellule une sorte de cadre rigide imperméable appelé cadre de Caspary du non de son découvreur. Ainsi pour traverser l’endoderme, l’eau et les sels minéraux ne peuvent qu’emprunter la voie du symplaste. Cette bande de Caspary empêche le reflux de l'eau et des sels minéraux du cylindre central vers la zone corticale
Document 12 : Traversée de l’eau au niveau de l’endoderme
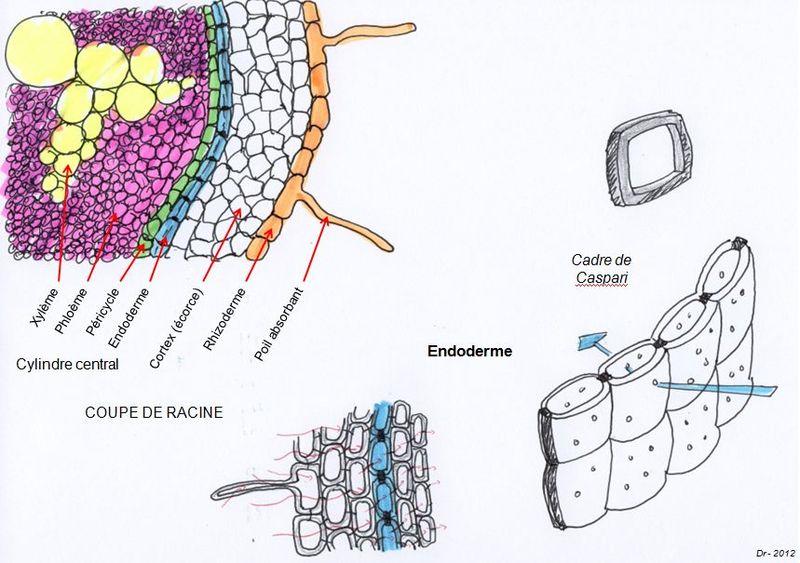
Source : Cadres de Caspary.JPG par David R, via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Cadres_de_Caspary.JPG
Passé l’endoderme, l’eau circule de nouveau par voie apoplasmique et se déverse dans les vaisseaux de xylème où elle constitue la sève brute contenant les sels minéraux et des acides aminés qui proviennent de la réduction des nitrates. Une fois déversée dans les vaisseaux, elle remonte par capillarité.
L’un des moteurs de l’ascension de l’eau et des sels minéraux est l’évapotranspiration des feuilles : la perte d’eau au niveau des stomates crée un « appel d’eau », un effet « aspirant ». En effet, puisqu'il n'y a que quelques traces de dioxyde de carbone dans l'air (0,037%), les stomates restent longtemps ouverts et la plante perd plusieurs centaines de molécules d'eau pour chaque molécule de CO2 qu'elle absorbe. Il y a d’abord évaporation de l’eau à partir des cellules qui bordent les chambres sous-stomatiques puis transpiration, c'est-à-dire diffusion de la vapeur d’eau par l’ostiole quand il est ouvert. La plante rejette ainsi dans l’atmosphère la presque totalité de l’eau qu’elle puise dans le sol.
L’autre moteur de la remontée d’eau est la pression racinaire. En sectionnant la tige d’une plante herbacée, on peut observer la sortie d’une goutte d’eau au niveau de la coupe sur la partie connectée aux racines. Cela s’observe d’autant mieux le matin : la nuit, les stomates sont fermés mais les poils absorbants continuent à absorber de l’eau. Or s’il y a beaucoup d'eau et qu'aucune eau n'est éliminée par transpiration, du fait de la présence du cadre de Caspary de l’endoderme, l’eau ne peut ressortir du cylindre central et elle s’y accumule: une pression hydrostatique positive apparaît, on parle de pression racinaire.
Même sans transpiration et pression racinaire, quand la plante est en équilibre hydrique avec le milieu extérieur, il existe un débit d'eau interne qui est tout à fait suffisant pour le transport des ions. Le cycle interne de l'eau dans le circuit montant et le circuit descendant est généré par l'eau du xylème qui s’écoulant vers le haut, finit par circuler dans le circuit opposé descendant : les deux systèmes sont interconnectés. Il ne faut pas imaginer une connexion des vaisseaux mais plutôt des vaisseaux qui se terminent en cul de sac et une transition de l’eau par les cellules constitutives des feuilles.
Les cellules du parenchyme situées entre l’épiderme inférieur et supérieur de la feuille récupèrent l’eau du xylème par voie apoplasmique. Elles sont également en contact avec les vaisseaux du phloème et y sont reliés par les plasmodesmes. Le chargement en sucres du tube criblé se fait donc par la voie symplasmique. L’eau du xylème va ainsi circuler à travers les cellules du parenchyme pour rejoindre les tubes criblés selon un gradient de pression osmotique. L’augmentation du taux de saccharose dans une cellule du tube criblé est responsable de ce gradient. L’arrivée d’eau dans cette cellule chargée en sucre provoque une augmentation de la pression hydrique et une circulation de l’eau chargée en sucres vers les cellules criblées situées en-dessous : le courant du phloème est ainsi créé. Le déchargement du saccharose au niveau des fruits, des graines, des différents organes de réserve ou des zones de croissance, se fait par la voie apoplasmique et symplasmique, toujours selon un gradient de concentration. Il provoque une baisse de la pression osmotique dans la cellule criblée subissant le déchargement ce qui provoque un transfert d’eau par osmose vers les tubes du xylème chargés en eau riche en sels minéraux. La boucle est bouclée…
Document 13 : La circulation interne de l’eau

Circulation de la matière - SVT - ENJEUX Term spé #2 - Mathrix
Date de dernière mise à jour : 24/12/2022