Les messages nerveux des réflexes
Vidéo en bas de page ^^
Les messages nerveux sont de nature électrique. Pour preuve, quand vous vous prenez « un coup de jus » vos muscles sont provisoirement immobilisés : plus aucun message ne passe.
I Les neurones présentent un potentiel de repos
Document 1 : Enregistrement d’un potentiel de repos sur l’axone de calmar géant.
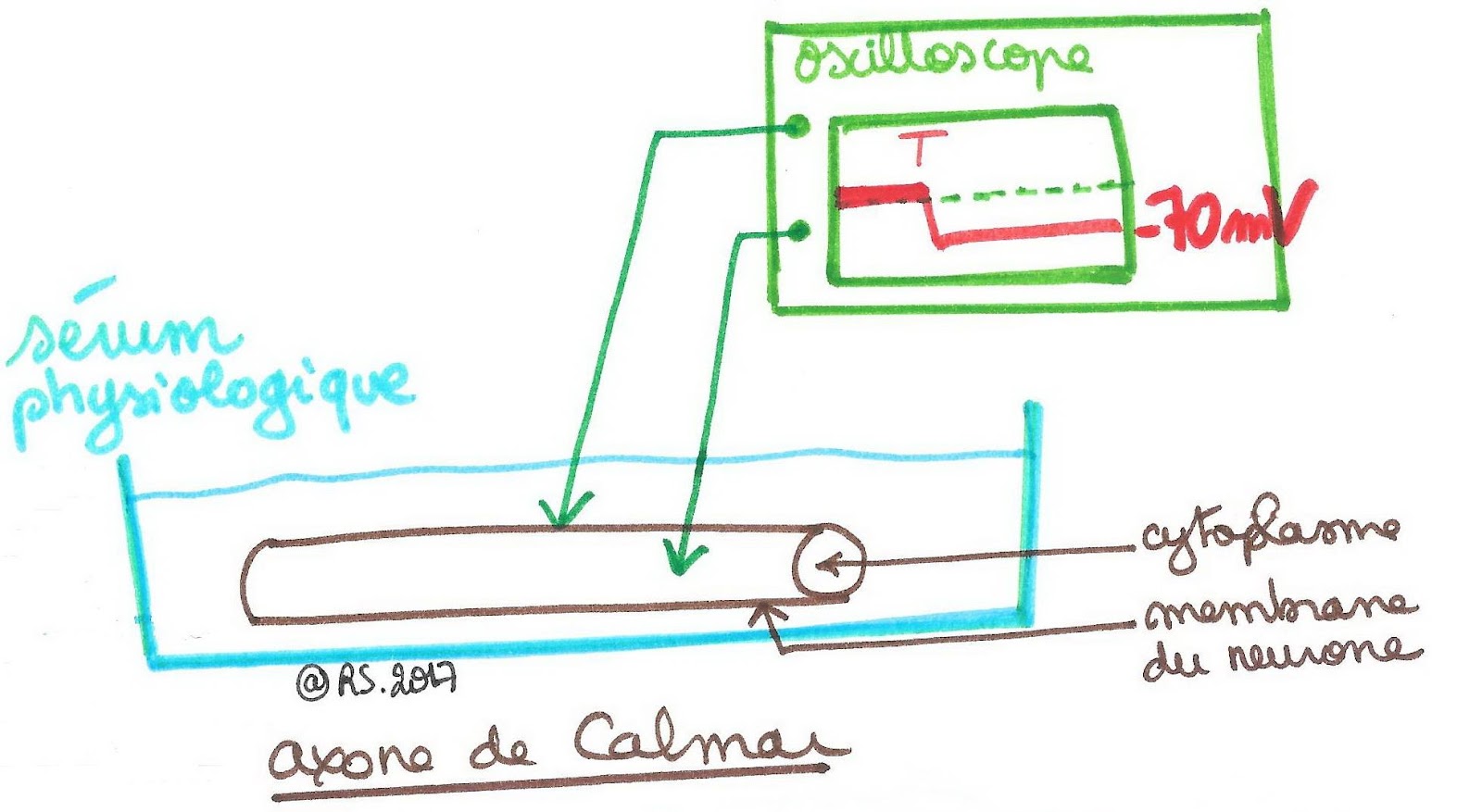
En absence de stimulation, la membrane de la fibre nerveuse est polarisée : la face interne est chargée plus négativement que la face externe. La membrane d’une cellule nerveuse au repos présente donc une différence de potentiel entre la face extracellulaire et la face intracellulaire (différence de potentiel transmembranaire ou ddp transmembranaire de - 70 mV en moyenne) : on parle de potentiel de repos. Toutes les cellules vivantes présentent cette ddp.
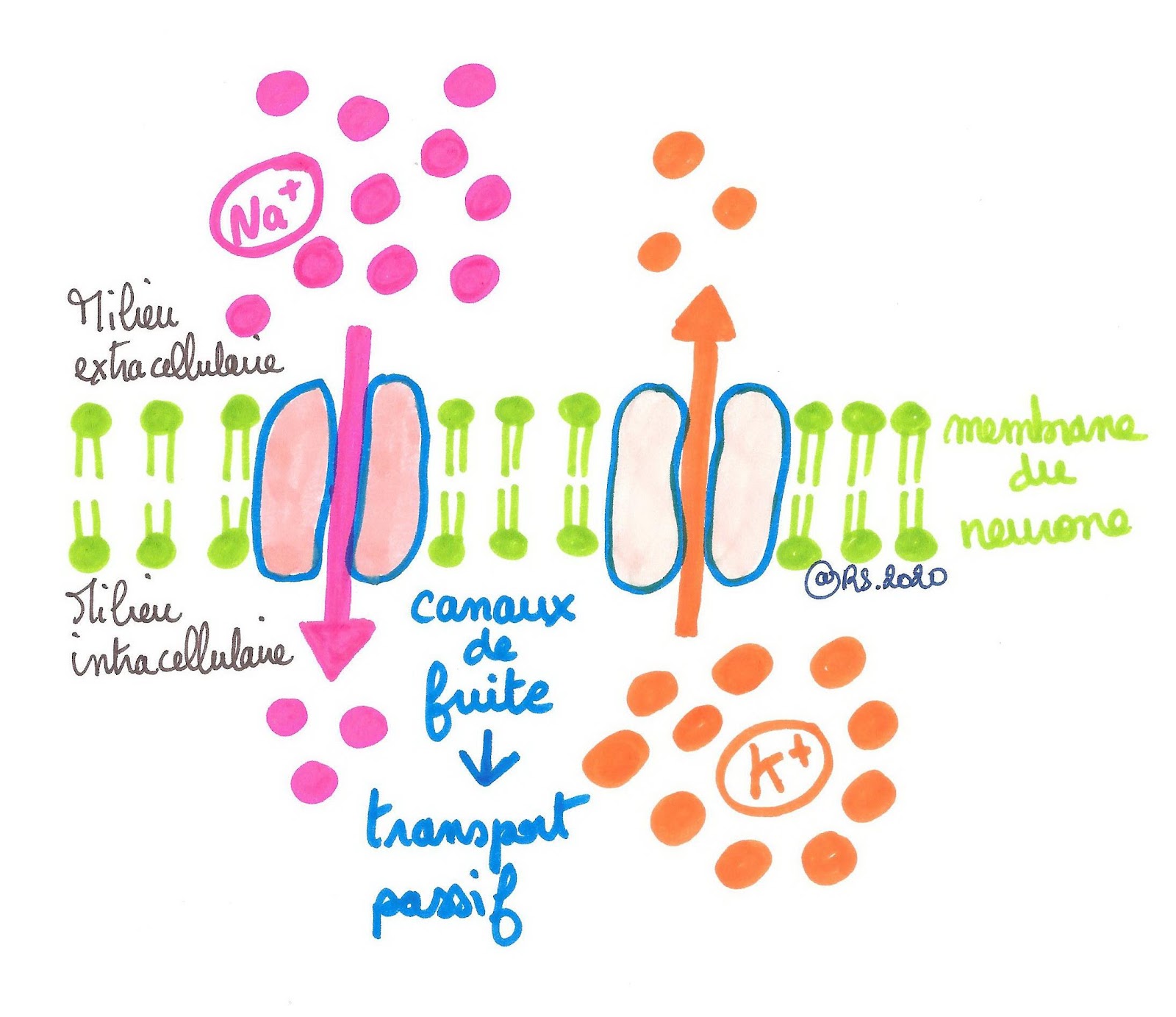
Au niveau de la membrane du neurone, il y a des protéines membranaires qui se comportent comme des canaux appelés canaux de fuite (ils sont toujours ouverts). À travers ces canaux, les ions sodium et potassium diffusent selon leur gradient de concentration (transport passif) :
• les ions Na+ diffusent du milieu extracellulaire (milieu le plus concentré) vers le milieu intracellulaire (le milieu le moins concentré).
• Les ions K+ diffusent du milieu intracellulaire (le milieu le plus concentré) vers le milieu extracellulaire (le milieu le moins concentré).
Document 2 : Diffusion passive des ions sodium et potassium au niveau de la membrane du neurone
Si on considère uniquement ce phénomène (transport passif), la diffusion de ces ions à travers la membrane aboutirait à une égalisation des concentrations intra et extracellulaires en potassium et sodium et par conséquent à l’annulation du potentiel de repos, ce qui n’est pas réalisable en situation physiologique normale. Le transport passif (ou diffusion) est insuffisant pour expliquer le déséquilibre ionique permanent de part et d’autre de la membrane.
Le potentiel de repos s’explique par la répartition inégale des ions sodium et potassium de part et d’autre de la membrane axonique. Cette répartition inégale des ions est un état d’équilibre déterminé par :
• un transport passif (diffusion) qui se fait selon le gradient des concentrations : entrée des ions sodium et sortie des ions potassium à travers des protéines membranaires qui se comportent comme des canaux de fuite toujours ouverts.
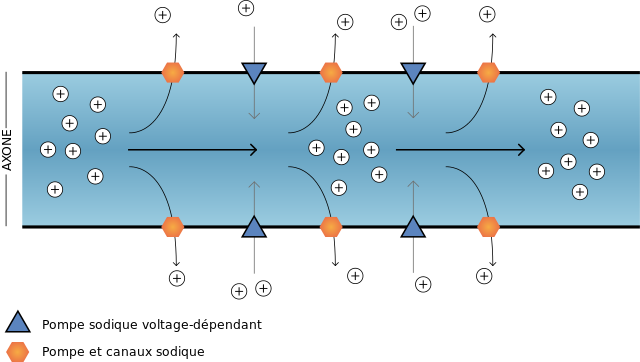
• Un transport actif qui se fait contre le gradient de concentration : sortie des ions sodium et entrée des ions potassium. Ce transport est assuré par une protéine membranaire qui joue le rôle d’une enzyme capable de catalyser l’hydrolyse de l’ATP. L’énergie libérée lors de l’hydrolyse de l’ATP est utilisée par cette même protéine pour assurer la sortie des ions Na+ et l’entrée des ions K+. Cette protéine membranaire jouant le rôle d’une enzyme est assimilée à une pompe Na-K-ATPase.
Document 3 : Pompe Na-K ATP ase et différence de potentiel transmembranaire

II La naissance du message nerveux sensitif
Document 4 : Enregistrement d’un potentiel d’action

La stimulation appliquée au niveau du fuseau neuromusculaire produit dans un premier temps un potentiel de récepteur qui est un potentiel local situé au niveau de l’extrémité de la fibre sensitive. Dès que l’amplitude de ce potentiel a atteint un "niveau seuil" au niveau du site générateur (-50 mV), il donne naissance à un potentiel dit potentiel d’action. L’énergie mécanique appliquée au niveau du fuseau neuromusculaire est convertie en énergie électrique : c’est la transduction. Au-dessus de cette valeur, lorsqu’on augmente l’intensité de la stimulation, les potentiels d’action enregistrés ont toujours la même amplitude. La réponse d’un neurone suit la loi du « tout ou rien ».
Document 5 : Loi du tout ou rien dans la réponse de la fibre nerveuse
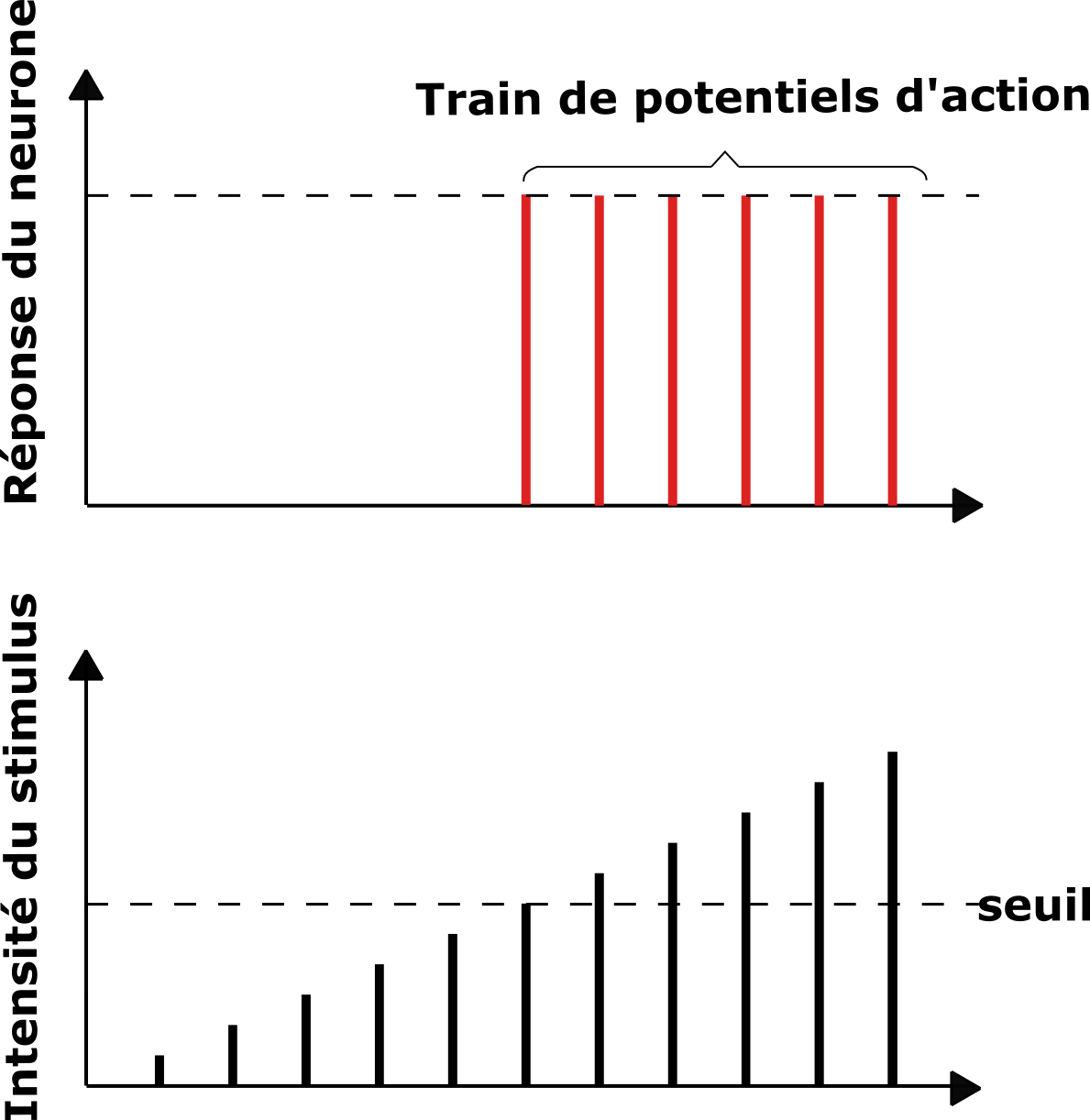
Loi du tout ou rien en.svg, par Blacktcvia Wikimedia commons, CC-BY-4.0, modifié par Sandra Rivière,
https://fr.wikibooks.org/wiki/Neurosciences/L%27influx_nerveux#/media/Fichier:All-or-none_law_en.svg
Le potentiel d’action correspond à un changement brutal et rapide (moins d’une milliseconde) de l’état d’équilibre ionique de la membrane du neurone. On observe dans un premier temps une inversion de la polarisation membranaire ou dépolarisation, puis une repolarisation. L’ensemble, de ces deux phases, dure en moyenne une milliseconde. Le potentiel d’action est suivi d’une phase où la négativité interne continue d’augmenter et dépasse la normale, il s’agit de l’hyperpolarisation consécutive au potentiel d’action.
1) La dépolarisation
À l’état de repos tous les canaux à sodium et potassium voltage-dépendants sont fermés. La stimulation provoque une légère dépolarisation de la membrane. Si la dépolarisation atteint le seuil de -50mV, les canaux à sodium voltage-dépendants s’ouvrent dans la zone dépolarisée. Par conséquent, on assiste à une augmentation considérable du flux entrant de sodium déjà existant. À cause de ces ions positifs entrants, l’intérieur de la cellule perd son électronégativité et devient même électropositive : le potentiel transmembranaire atteint + 30 mV. Le milieu extérieur auparavant électropositif devient naturellement électronégatif car il perd des charges positives. C’est la dépolarisation de la membrane.
Document 6 : Mécanismes ioniques de la dépolarisation

2) La repolarisation
La perméabilité de la membrane au potassium augmente car les canaux voltage-dépendants à sodium se ferment rapidement (au bout d’1 ms) et les canaux voltage-dépendants à potassium s’ouvrent. La fuite de potassium en dehors de la cellule est intensifiée pour tenter de rééquilibrer les charges électriques. Les ions K+ sortent donc de la cellule et rejoignent le milieu extérieur. Le cytoplasme perd des charges positives potassiques et a donc tendance à redevenir électronégatif. Pendant ce temps, le milieu extérieur gagne des ions positifs et a donc tendance à perdre son électronégativité. Il redevient électropositif. C’est la phase de repolarisation rapide.
Document 7 : Mécanismes ioniques de la repolarisation

3) L’hyperpolarisation.
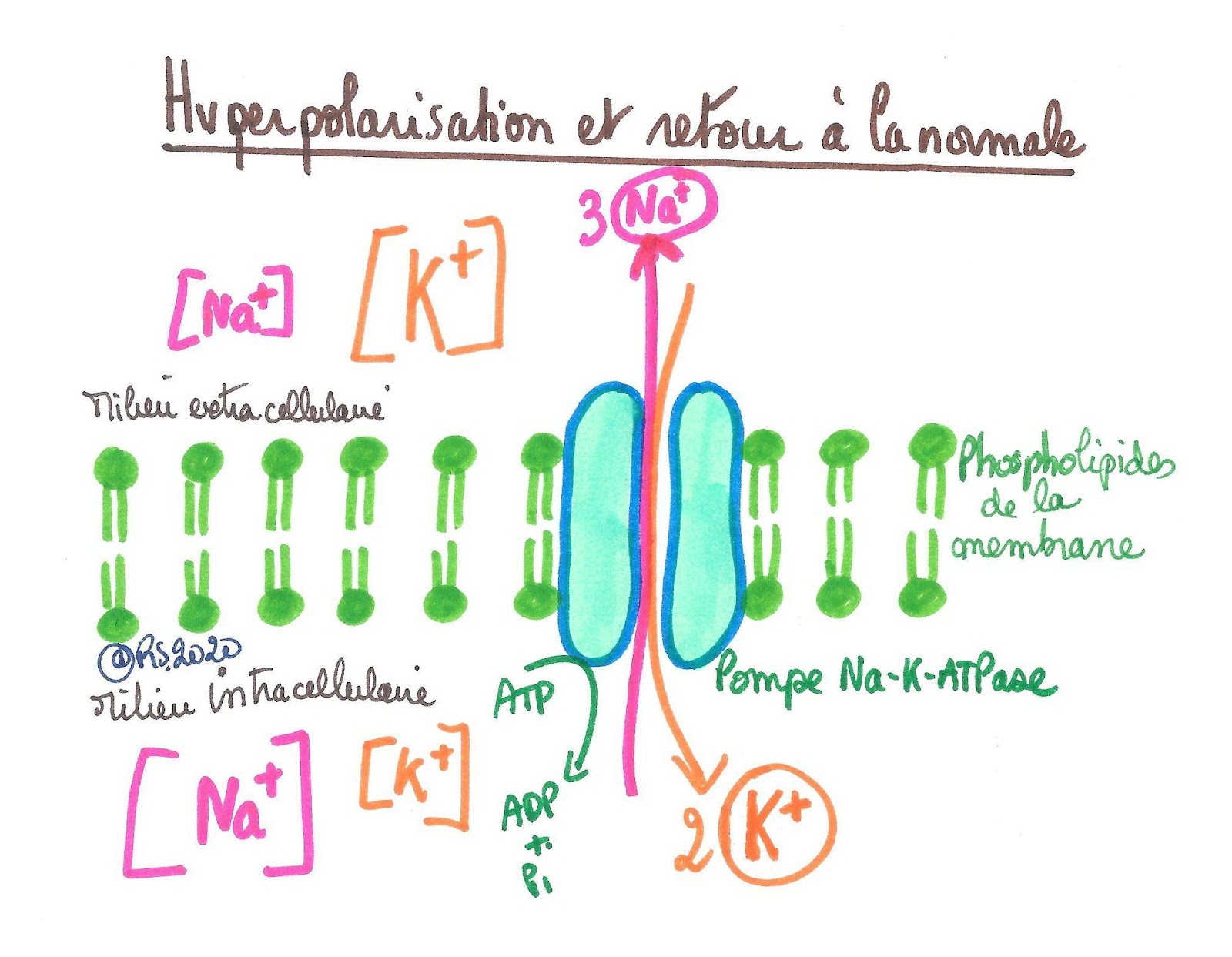
Les canaux à potassium restent ouverts et les canaux à sodium restent fermés. Le potentiel d'équilibre du potassium étant plus négatif (-80 mV) que le potentiel de repos (-65 mV), le potentiel de membrane devient plus négatif que le potentiel de repos : on parle d'hyperpolarisation.
La sortie des ions K+ par ces canaux vient donc contrecarrer toute dépolarisation ultérieure de la cellule : le potentiel de membrane est hyperpolarisé tant que les canaux potassiques restent ouverts. Aussi faut-il plus de courant dépolarisant pour que le potentiel de membrane atteigne à nouveau le seuil de dépolarisation : c'est la période réfractaire.
Il y a finalement un retour progressif au potentiel de repos, c’est-à-dire un retour aux conditions électriques et de concentrations ioniques antérieures : on observe une normalisation de la perméabilité de la membrane et une mise en jeu intensifiée des pompes Na-K ATPase pour ramener les concentrations ioniques aux valeurs antérieures. Il y a rejet accru de sodium à l’extérieur de la cellule et récupération accrue de potassium à l’intérieur de la cellule.
Document 8 : État d’hyperpolarisation et mécanisme ionique de retour au potentiel de repos
La création du potentiel de repos est donc le pré-requis pour qu’une cellule excitable génère un nouveau potentiel d’action.
III La propagation du message nerveux
1) Le codage du message nerveux
Tous les potentiels d'action ayant la même amplitude, le codage de l'influx nerveux se fait donc en modulation de fréquence : lorsque l’on augmente l’intensité de stimulation, la fréquence des potentiels d’action est également en augmentation. Ainsi au niveau d’une fibre nerveuse, le message est codé en fréquence de potentiels d’action.
Document 9 : Illustration du codage en fréquence du potentiel d’action
On soumet un neurone à un stimulus initialement peu intense, puis moyennement intense et enfin très intense. On voit que la fréquence d'émission des potentiels d'action augmente de plus en plus avec l'intensité du stimulus.

NeuralMassSimulation.png, par via Wikimedia commons, CC-BY-3.0, modifié par Sandra Rivière, https://commons.wikimedia.org/wiki/File:NeuralMassSimulation.png
Au niveau d’un nerf, on peut mesurer un potentiel global, dû à l’ensemble des potentiels d’actions qui circulent dans les fibres nerveuses. L’amplitude du potentiel global augmente avec l’intensité de stimulation : il y a un recrutement plus important de fibres nerveuses. Au niveau d’un nerf, le message nerveux est codé en amplitude de potentiel global.
Document 10 : Enregistrement du potentiel global d’un nerf stimulé

2) Un conduction de proche en proche
L'influx se déplace le long de l'axone dans une seule direction, vers la terminaison axonale où il y a relais avec d'autres neurones. On dit qu’il se propage de proche en proche.
Document 11 : Propagation du potentiel d’action de proche en proche dans l’axone
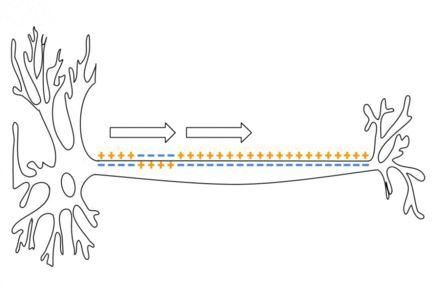
Action Potential.gif,par Laurentaylorj via wikimedia commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Action_Potential.gif
Lorsqu'un potentiel d'action apparaît à un endroit donné de l'axone, la portion voisine qui lui a donné naissance entre en période réfractaire, ce qui l'empêche d'être excitée à son tour. Cette période réfractaire est expliquée par la désensibilisation des canaux sodiques voltage-dépendants.
En revanche, la portion voisine qui n'a pas encore présenté de potentiel d'action commence à être excitée. Cette excitation provient de petits courants électriques très locaux qui s'établissent entre portion excitée et portion non encore excitée. Si la portion voisine est suffisamment dépolarisée (si elle atteint le seuil) un potentiel d’action sera créé. De proche en proche, se créent donc les conditions de naissance d'un potentiel d'action à côté de la portion qui est en train de réaliser un potentiel d'action (propagation régénérative).
Ainsi, la période réfractaire explique l'unidirectionalité de l'influx nerveux, depuis le cône d'émergence jusqu'à ses extrémités, les terminaisons synaptiques.
L'influx nerveux conserve toutes ses caractéristiques (amplitude, fréquence) durant sa progression : il est conservatif.
Document 12 : Principe de la dépolarisation de proche en proche
Propagation influx nerveux proche en proche.svg par fr: Utilisateur: Slagt via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Propagation_influx_nerveux_proche_en_proche.svg
Document 13 : Modélisation en étapes de la dépolarisation de proche en proche
Image 1 : Membrane du neurone à l’état de repos
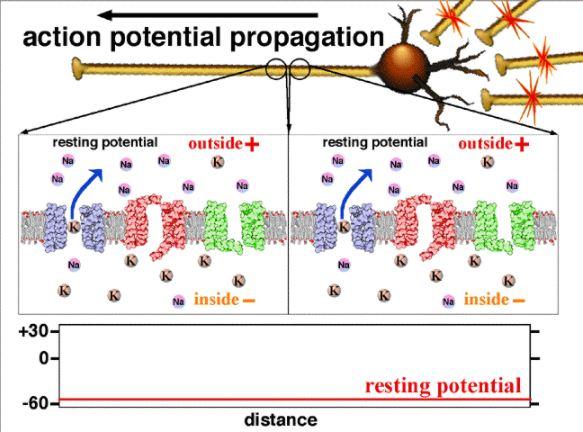
Image 2 : Dépolarisation suite à une stimulation, seuil atteint et naissance d’un potentiel d’action dans la première zone
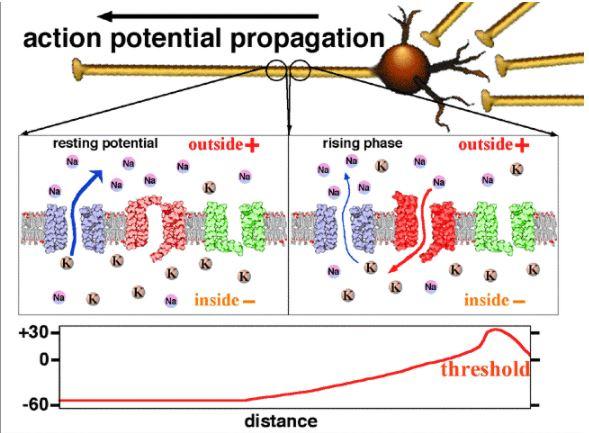
Image 3 : Ouverture des canaux voltages-dépendants voisins et début de dépolarisation de la zone voisine (à gauche)
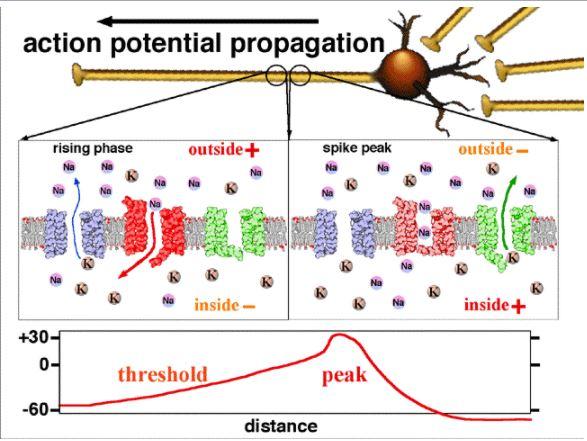
Image 4 : Naissance d’un potentiel d‘action dans la zone voisine, état réfractaire (hyperpolarisation) dans la première zone
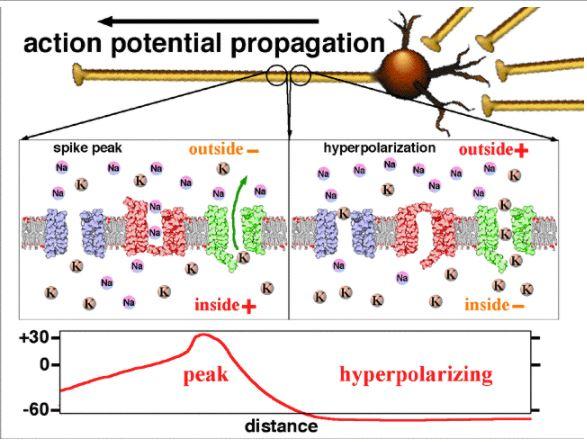
Image 5 : Fermeture des canaux voltage-dépendants de la deuxième zone, première zone en cours de rééquilibrage ionique.

Image 6 : État réfractaire de la deuxième zone, état de repos pour la première zone qui peut à nouveau produire un potentiel d’action.

Action potential propagation animation.gif, par John Schmidt via Wikimedia commons, CC-BY-SA-4.0,3.0,2.5,2.0,1.0, https://commons.wikimedia.org/wiki/File:Action_potential_propagation_animation.gif
3) La transmission saltatoire
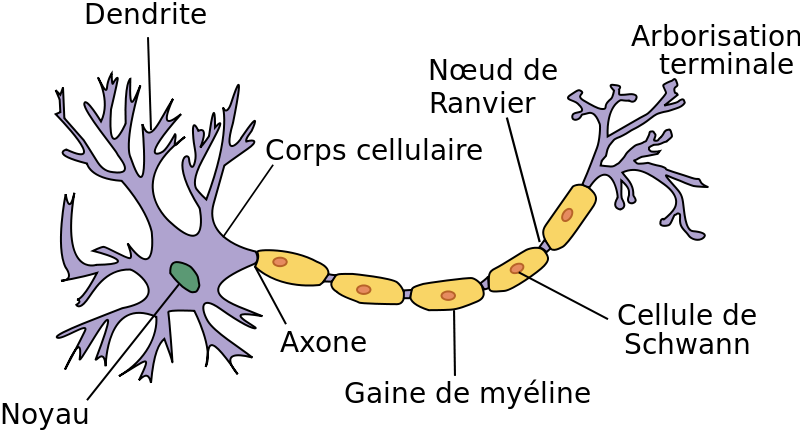
La conduction peut se faire soit de proche en proche le long de l'axone lorsque ce dernier est nu, soit de manière saltatoire lorsque l'axone possède une gaine de myéline. La myéline est maintenue autour de l'axone par des cellules appelées cellules de Schwann pour les neurones du système nerveux périphérique (ensemble des nerfs).
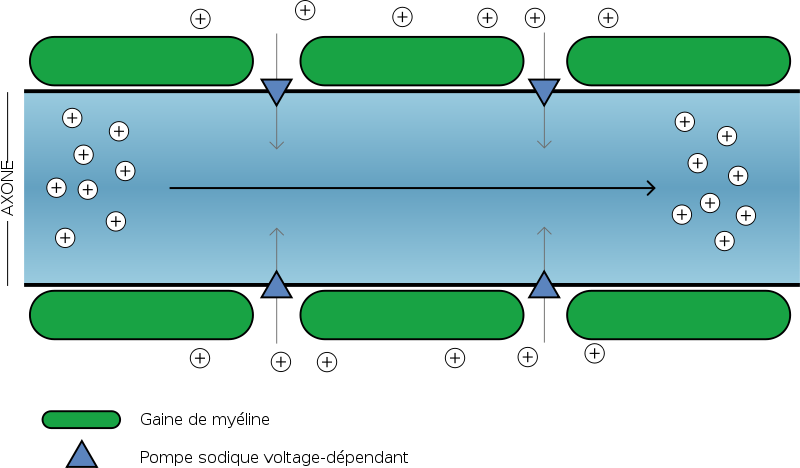
Chacune de ces cellules protectrices est séparée de ses deux voisines par un petit espace appelé nœud de Ranvier. La gaine de myéline joue le rôle d’isolant électrique : l’influx nerveux « saute » alors de nœud de Ranvier en nœud de Ranvier, ce qui permet une conduction beaucoup plus rapide (jusqu'à plus de 100 m/s, au lieu d'environ 10 m/s pour une fibre non myélinisée). Cette propagation en bond (ou en saut) donne son nom à la conduction saltatoire.
Document 14 : Neurone présentant une gaine de myéline
Neurone français schéma.svg, Aucun auteur lisible par machine fourni. Trame supposée (basée sur les revendications de droits d'auteur), via Wikimédia Commons, CC-BY-SA-3.0-migré, https://commons.wikimedia.org/wiki/File:Neurone_fran%C3%A7ais_sch%C3%A9ma.svg
Les charges positives (Sodium) se déplacent au bout de l'axone suivant le gradient électrochimique. Les gaines de myélines isolent l'axone, rien ne sort, rien ne rentre à leur niveau. Entre deux gaines (nœud de Ranvier), on retrouve des pompes sodiques voltage-dépendantes régénérant le potentiel d'action initial.
Document 15 : Mécanisme de la conduction saltatoire
Propagation influx nerveux axone myélinisé.svg par Utilisateur: Slagt via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Propagation_influx_nerveux_axone_my%C3%A9linis%C3%A9.svg
IV La transmission du message nerveux
Les neurones sont organisés en réseau ce qui implique qu’ils sont connectés. Le message doit donc passer d’un neurone à un autre. Cela signifie que l’arrivée d’un train de potentiels d’action à l’extrémité d’un axone doit déclencher des potentiels d’action sur le neurone suivant. La zone de contact entre 2 neurones est appelée synapse.
La synapse (du grec syn = ensemble et haptein = toucher, saisir) désigne une zone de contact qui s'établit entre deux neurones (synapse neuro-neuronique), ou entre un neurone et une autre cellule comme la synapse neuromusculaire.
1) La synapse neuro-neuronique entre le neurone sensitif et le motoneurone alpha
La plupart des synapses ne présentent pas de contact physique entre les deux cellules : une fente, appelée fente synaptique, empêche toute communication de nature électrique. Un messager doit prendre le relais entre les deux neurones impliqués : celui-ci est de nature chimique.
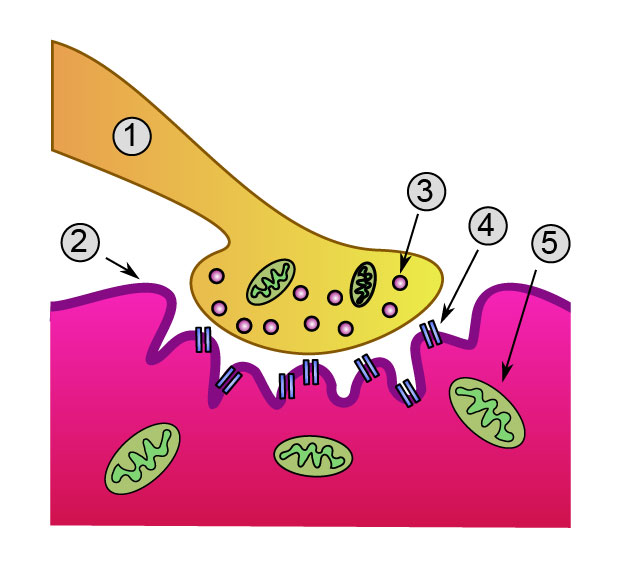
Document 16 : Fonctionnement d’une synapse neuro-neuronique
Transmission chimique du neurone A (présynaptique) au neurone B (postsynaptique)
1. Mitochondrie permettant l’apport énergétique nécessaire à la synthèse du neurotransmetteur
2. Vésicule synaptique contenant des molécules de neurotransmetteur
3. Autorécepteur membranaire
4. Fente synaptique avec neurotransmetteur libéré (ex : sérotonine ou dopamine)
5. Récepteurs postsynaptiques activés par le neurotransmetteur (induction d'un potentiel postsynaptique)
6. Canal calcium
7. Exocytose d'une vésicule
8. Neurotransmetteur recapturé

Synapse diag1.svg par vectorisation: Mouagip ( discussion ), Synapse_diag1.png : Dessiné par fr: Utilisateur: Dake, Corrections du PNG original par fr: User: Nrets, via Wikimédia Commons, CC-BY-SA-3.0-migré, https://commons.wikimedia.org/wiki/File:Synapse_diag1.svg
Au niveau de la synapse, la transmission du message nerveux est unidirectionnelle. La dépolarisation d'une terminaison synaptique par un potentiel d'action provoque l'ouverture de canaux perméables au calcium et l'entrée de cet ion dans la terminaison axonique. Sous l'impulsion de cet influx de calcium, les vésicules fusionnent avec la membrane cellulaire et libèrent leur contenu dans le milieu extracellulaire : on parle d’exocytose. Le neurotransmetteur, dans ce cas de l’acétylcholine, va alors diffuser librement et atteindre la membrane de la cellule cible à laquelle la terminaison nerveuse est associée. Le délai est de 0.6 à 0.8 millisecondes. La première étape de la transmission revient donc à convertir un signal électrique (le potentiel d'action) en signal chimique (le neurotransmetteur).
La cellule cible possède sur sa membrane des protéines réceptrices auxquelles des molécules de neurotransmetteur viennent s'associer spécifiquement, comme une clé dans une serrure. La liaison du neurotransmetteur à ces récepteurs dits ionotropes va généralement provoquer l'ouverture (ou plus rarement la fermeture) de ceux-ci : selon le type d'ions auxquels ils sont perméables, il en résulte une dépolarisation ou une hyperpolarisation de la cellule cible. Cette deuxième étape de la transmission synaptique revient donc à convertir un signal chimique en un signal électrique.
Document 17 : Fonctionnement d’un récepteur ionotrope

Ainsi au niveau de la synapse, selon la quantité des neurotransmetteurs, un message postsynaptique prend naissance ou pas. Le message est codé en quantité de neurotransmetteur libéré : plus la fréquence des potentiels d’action présynaptiques est élevée, plus la quantité de neurotransmetteur libéré est importante.
Document 18 : Relation entre fréquence des potentiels d’action présynaptiques et la quantité de neurotransmetteurs libérée
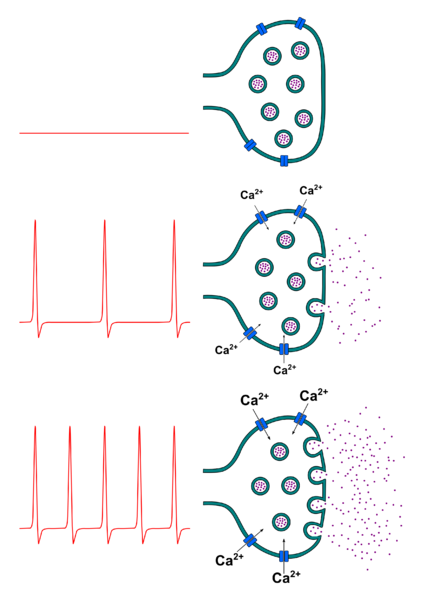
Neurotransmitter release.png par Peter Duncan propre travail, via Wikimedia commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Neurotransmitter_release.png
Pour éviter que la stimulation du neurone postsynaptique ne se prolonge, deux systèmes éliminent la molécule de l'espace inter-synaptique :
- la dégradation, qui met en jeu des enzymes spécifiques qui vont métaboliser le neurotransmetteur, mettant fin à son effet sur le neurone postsynaptique
- la recapture, pendant laquelle le neurotransmetteur( ou ses précurseurs issus de la dégradation enzymatique) est récupéré par le neurone présynaptique pour être réutilisé ou détruit.
Document 19 : Mécanisme d’arrêt du fonctionnement synaptique

Generic_Neurotransmitter_System, par NIDA (NIH) via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Generic_Neurotransmitter_System.jpg
2) La synapse neuromusculaire du motoneurone alpha
Le motoneurone répond à la libération du neurotransmetteur de la fibre afférente par la naissance d’un message nerveux moteur efférent conduisant à la contraction du muscle étiré. Lorsqu’un message nerveux est conduit le long de l’axone du motoneurone, il en résulte l’exocytose ou la libération de molécules d’un neurotransmetteur, l’acétylcholine, au niveau des boutons synaptiques formant les terminaisons axonales.
Document 20 : Structure de la synapse neuromusculaire
Jonction neuromusculaire (vue rapprochée)
- Axone présynaptique
- Sarcolemme (membrane plasmique de la fibre musculaire)
- Vésicules synaptiques
- Récepteurs de l'acétylcholine
- Mitochondrie
Synapse_diag4, Dessiné par Dake avec Inkscape 0.42., Corrigé par en: User: Nrets, via Wikimédia Commons, https://commons.wikimedia.org/wiki/File:Synapse_diag4.png
Après diffusion du neurotransmetteur dans l’espace synaptique séparant la terminaison axonale du muscle au niveau de la jonction neuromusculaire, les molécules de neurotransmetteur se lient à des récepteurs spécifiques présents dans la membrane des fibres musculaires. La formation de complexes neurotransmetteur-récepteur provoque une dépolarisation de la membrane qui, si elle atteint le seuil, déclenche la contraction du muscle On parle de synapse neuromusculaire. Un ensemble de synapses neuromusculaires est appelé plaque motrice. C’est la concentration en neurotransmetteur qui constitue le code du message : plus la concentration en acétylcholine libérée est importante plus la fréquence des potentiels d’action musculaires sera importante, provoquant une contraction musculaire de plus forte amplitude. Afin de contrôler la contraction du muscle, l’acétylcholine est rapidement détruite par l’acétylcholinestérase et permet un bon fonctionnement des muscles.
Document 21 : Fonctionnement de la synapse neuromusculaire
{kind=link}
{kind=link}
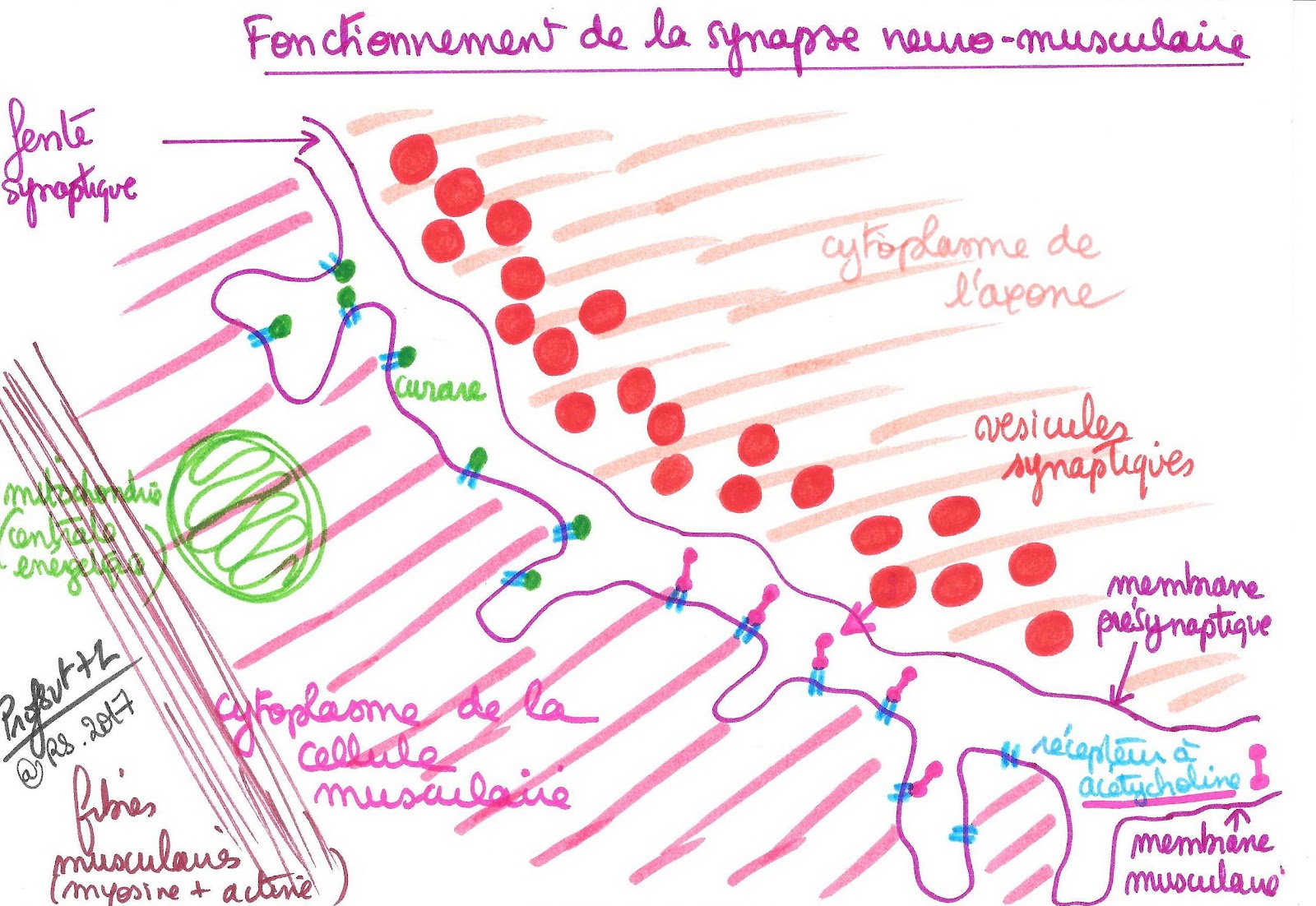
Certaines substances sont capables de modifier le fonctionnement de la synapse en se fixant sur les récepteurs de la membrane du neurone post-synaptique.
Le curare (utilisé dans les myorelaxants) se fixe à la place de l’acétylcholine et bloque les synapses neuromusculaires en prenant la place de l’acétylcholine qui ne peut alors plus agir. Il provoque un relâchement durable des muscles qui est mortel par paralysie des muscles respiratoires. C’est un antagoniste de l’acétylcholine car son action est inverse à celle-ci. Il existe aussi des substances qui empêchent l’élimination de l’acétylcholine dans la fente synaptique : elles prolongent la durée d’action du neurotransmetteur (gaz moutarde…)
Les messages nerveux du réflexe- SVT - SANTÉ Term spé #2 - Mathrix
Date de dernière mise à jour : 20/06/2021