Fleurs et pollinisation
Vidéo en bas de page ^^
I Rappels et contexte d’étude
Il existe deux grandes catégories de cycles de vie :
- les cycles de vie courts réalisés sur une ou deux années, on parle alors de plantes annuelles ou bisannuelles,
- les cycles de vie longs réalisés sur plusieurs années, on parle de plantes vivaces.
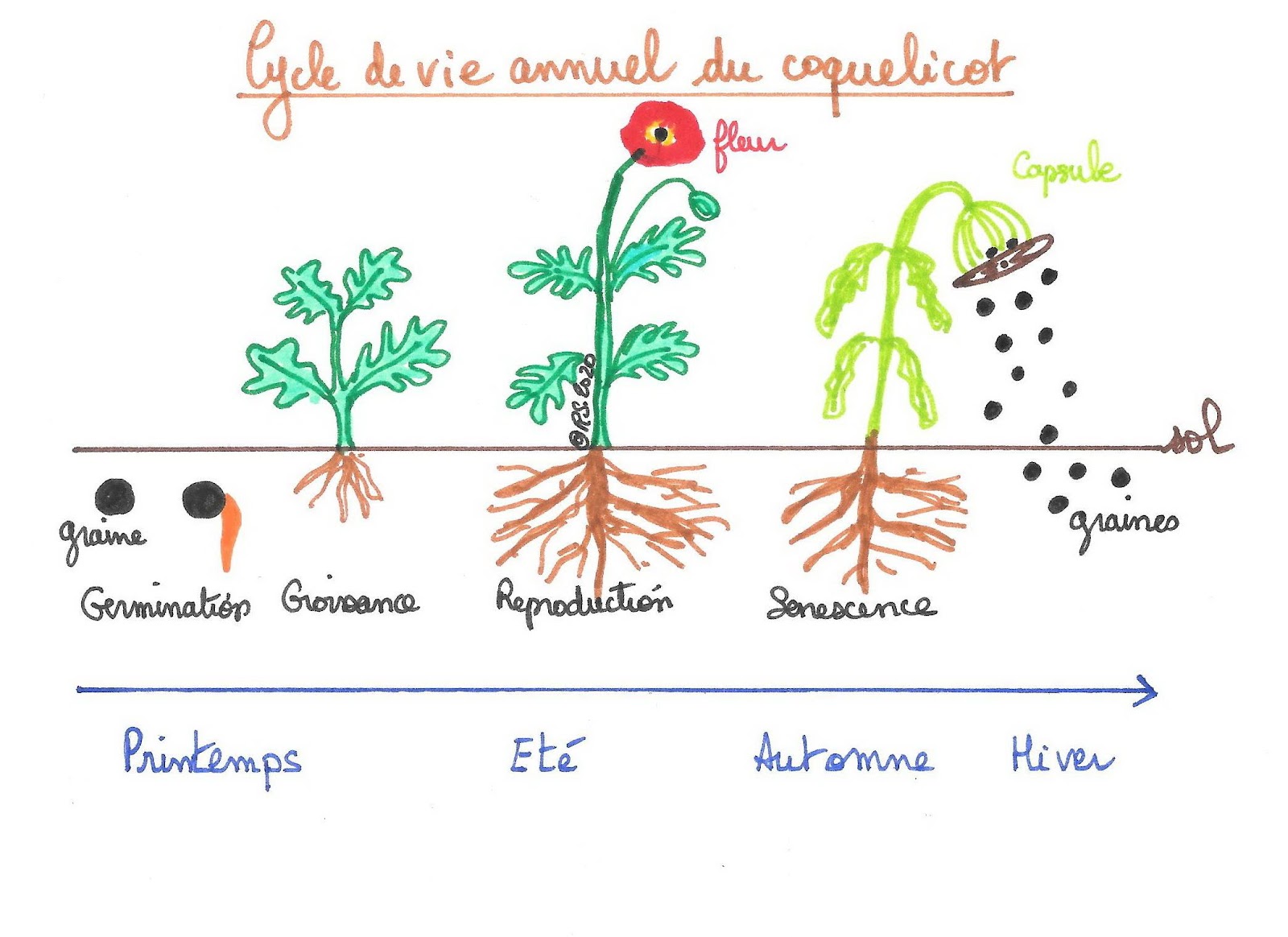
Document 1 : Cycle de vie d’une plante annuelle : le coquelicot
Document 2 : Cycle de vie d’une plante bisannuelle : la carotte

Document 3 : Cycle de vie d’une plante vivace : le chêne
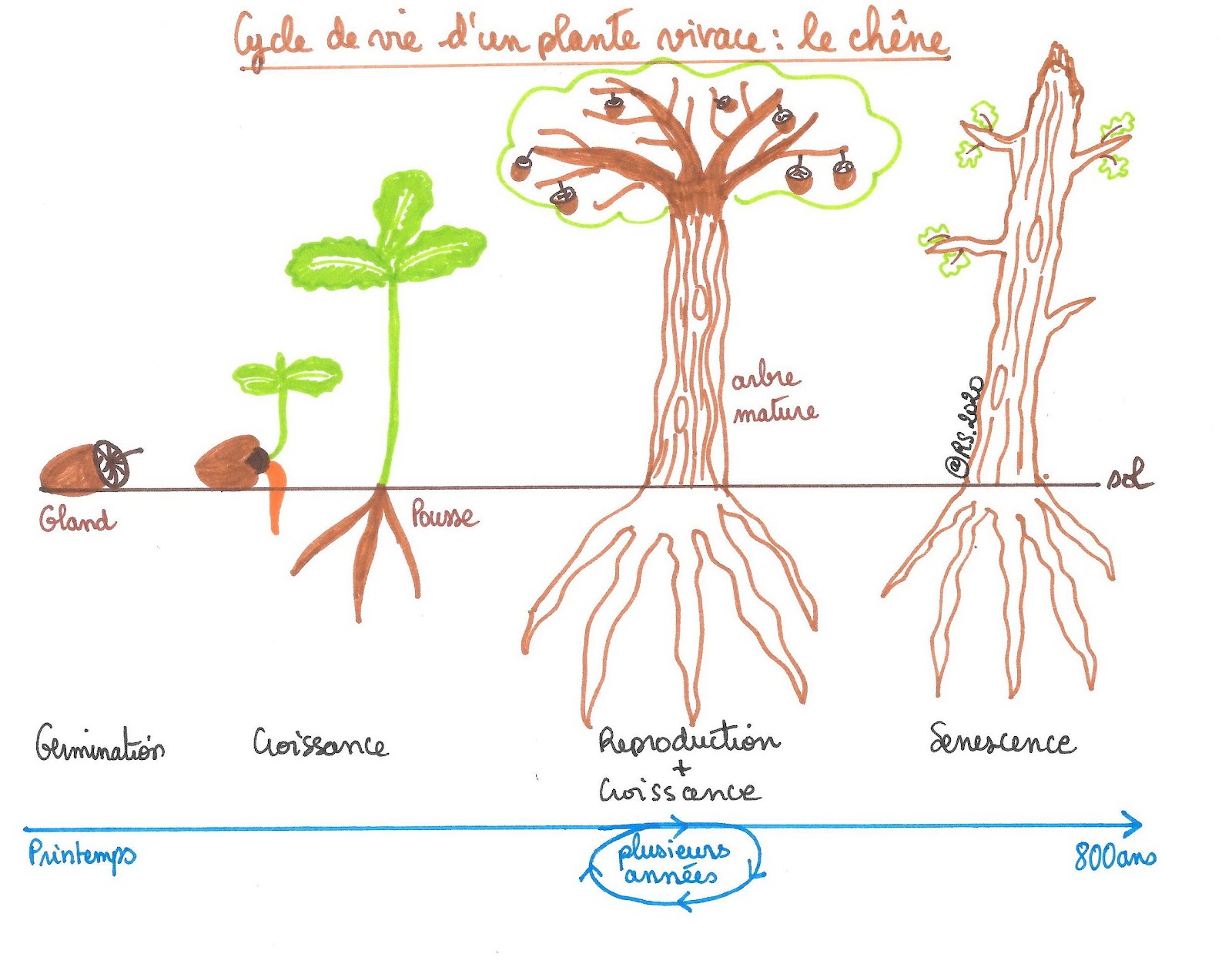
Dans les deux cas, le cycle de vie d’une plante à fleurs est marqué par de grandes étapes : la croissance, une vie adulte ponctuée de périodes de reproduction et une sénescence conduisant à la mort. Quel que soit le type de plantes, ces dernières ont une vie soumise à des contraintes liées à la vie fixée comme trouver leurs ressources alimentaires dans leur milieu de vie ou encore résister aux différents changements de saisons.
Document 4 : Tableau répertoriant les contraintes de plantes à vie fixée et leurs conséquences
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Les plantes terrestres montrent une capacité d’adaptation à la vie fixée aux interfaces sol/atmosphère ou sol/hydrosphère tant au niveau de leur structure que de leur fonctionnement.
Le système racinaire d’une plante offre une grande surface d’échanges avec le sol permettant l’alimentation en eau et en sels minéraux, ainsi que l’ancrage de la plante. Le système caulinaire avec sa surface foliaire offre une grande surface de captation de l’énergie lumineuse également impliquée dans les échanges gazeux permettant la photosynthèse. Ces deux systèmes assurent les deux grandes fonctions que sont la nutrition et la croissance.
Les variations des paramètres de l’environnement sont détectées par les plantes qui en réponse, modifient certaines de leurs caractéristiques structurales comme la perte des feuilles ou le développement des bourgeons et des fleurs.
Ainsi l’adaptation aux fluctuations des paramètres de l’environnement induit des modifications des grandes fonctions à l’origine des cycles de vie.
II La fleur, un appareil reproducteur
Il existe des plantes qui produisent des graines nues non protégées par des fruits : ce sont les gymnospermes dont les plus connus sont les sapins.
Certaines plantes développent des organes spécialisés dans la production de graines protégées par des fruits : les fleurs. C’est le groupe des plantes à fleurs ou Angiospermes.
Il existe des fleurs sans pétales et des fleurs avec pétales. Les fleurs sans pétales possèdent des feuilles modifiées rigides appelées glumes et glumelles qui enserrent les organes reproducteurs et les protègent. Ce sont typiquement les fleurs de graminées.
Document 5 : Structure d’une fleur de graminée

Source : Urochloa mosambicensis spikelet12 annoté - Flickr - Macleay Grass Man.jpg par Harry Rose de Dungog, Australie, via Wikimédia commons, CC-BY-2.0, https://commons.wikimedia.org/wiki/File:Urochloa_mosambicensis_spikelet12_annotated_-_Flickr_-_Macleay_Grass_Man.jpg
On observe soit des fleurs solitaires soit des inflorescences, c'est-à-dire des groupes de fleurs. Dans ce cas soit l’inflorescence est définie quand la première fleur à s’ouvrir est au sommet de la tige bloquant ainsi la croissance de la tige, soit l'inflorescence est indéfinie quand les fleurs les plus anciennes sont en dessous du sommet de la tige, celui-ci étant occupé par un bourgeon floral qui n’empêche pas la tige de grandir.
La plupart des fleurs sont hermaphrodites car elles possèdent à la fois un appareil reproducteur mâle l’androcée et un appareil reproducteur femelle le gynécée. Les fleurs peuvent présenter une symétrie bilatérale ou une symétrie radiale. On parle alors soit de fleurs zygomorphes soit de fleurs actinomorphes.
Document 6 : Symétries de la fleur
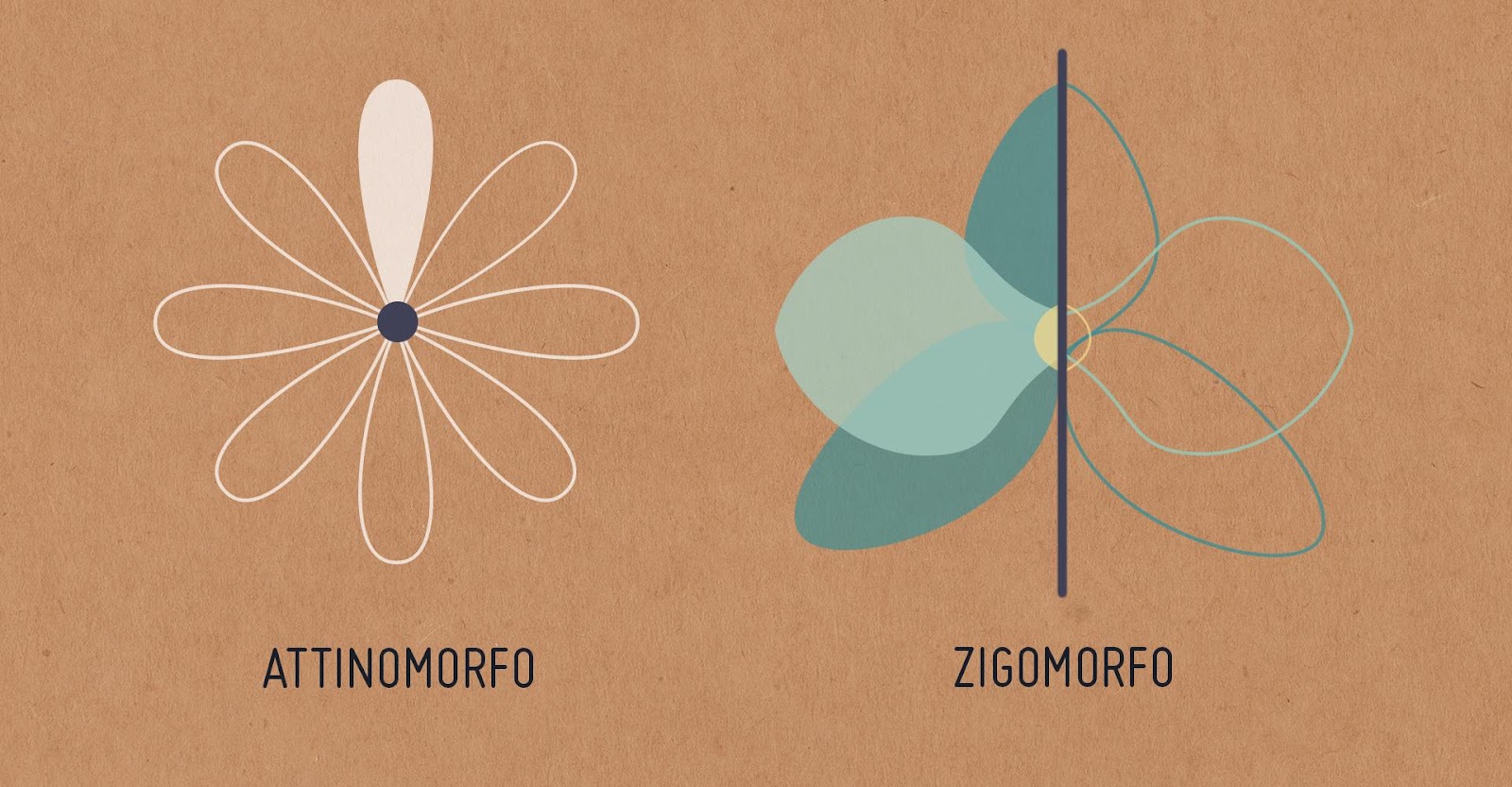
Attinomorfo e Zigomorfo.jpg, par Borgetti N., Isocrono D. (DISAFA), via Wikimédia Commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Attinomorfo_e_Zigomorfo.jpg
Une fleur avec pétales est portée par un pédoncule floral. Le sommet du pédoncule floral est renflé en un réceptacle floral qui porte des feuilles modifiées appelées pièces florales. Elles n’ont pas toutes la même fonction : certaines sont fertiles et d’autres stériles. Ces pièces sont disposées, de l’extérieur vers l’intérieur, sur 4 cercles concentriques appelés verticilles.
Document 7 : Structure de la fleur
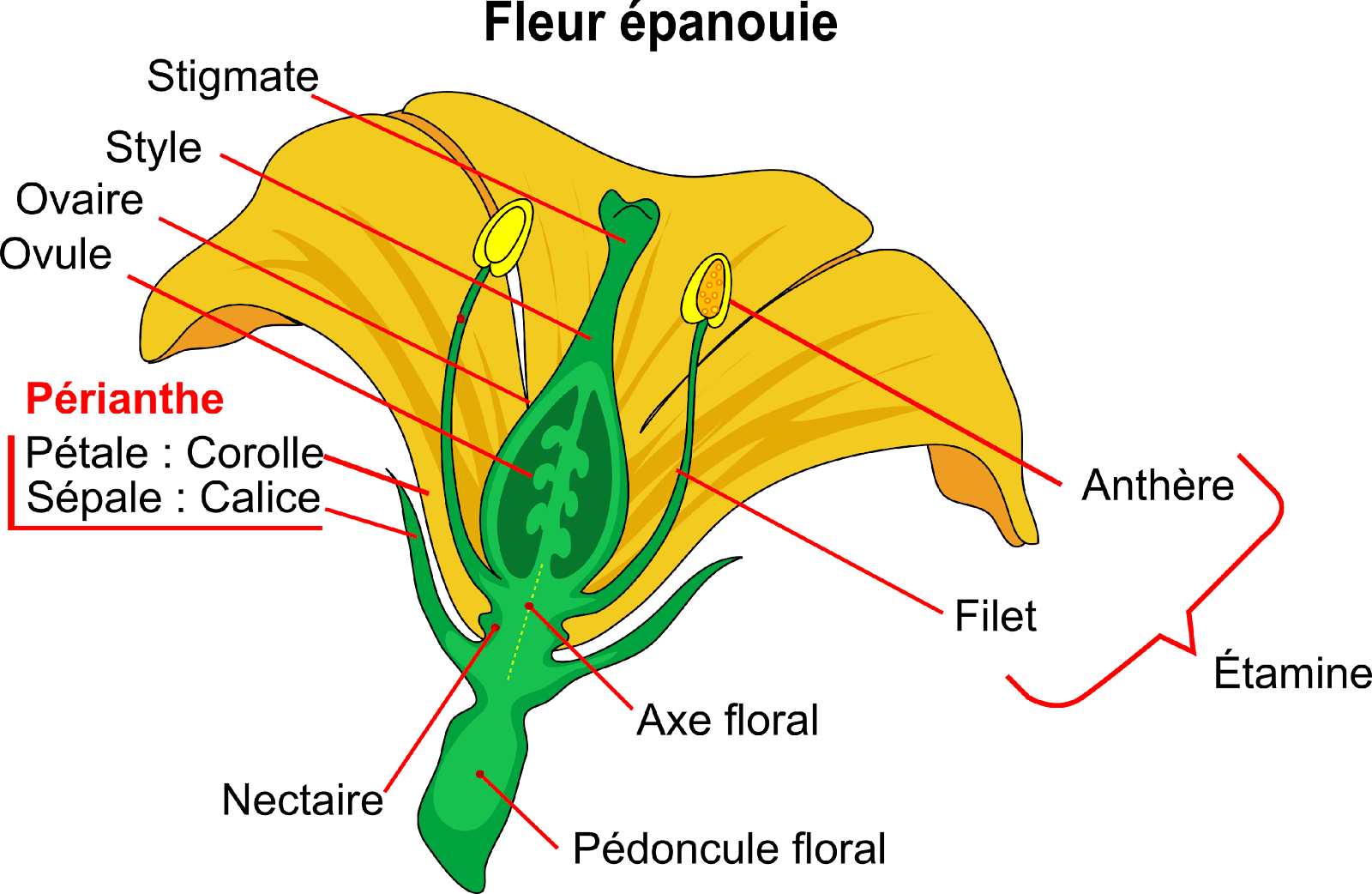
Mature flower diagram-fr.svg, par Mariana Ruiz LadyofHats, translation by Berrucomons ; via Wikimédia Commons, domaine publique, modifié par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Mature_flower_diagram-fr.svg?uselang=fr
Sur le verticille le plus extérieur, appelé le calice, on trouve les sépales qui protègent la fleur en bouton et qui s’écartent quand la fleur s’épanouie (s’ouvre). Les sépales quand ils sont colorés et de grandes tailles peuvent participer à l’attraction des pollinisateurs.
Disposés sur le verticille suivant, la corolle, les pétales qui jouent à la fois un rôle protecteur pour les pièces centrales fertiles et un rôle attractif pour les animaux pollinisateurs. L’ensemble calice et corolle forme ce qu’on appelle le périanthe. Quand les sépales et les pétales ne sont pas distinguables, on parle de tépales comme chez le lys. À la base des pétales on peut observer des glandes nectarifères appelées nectaires.
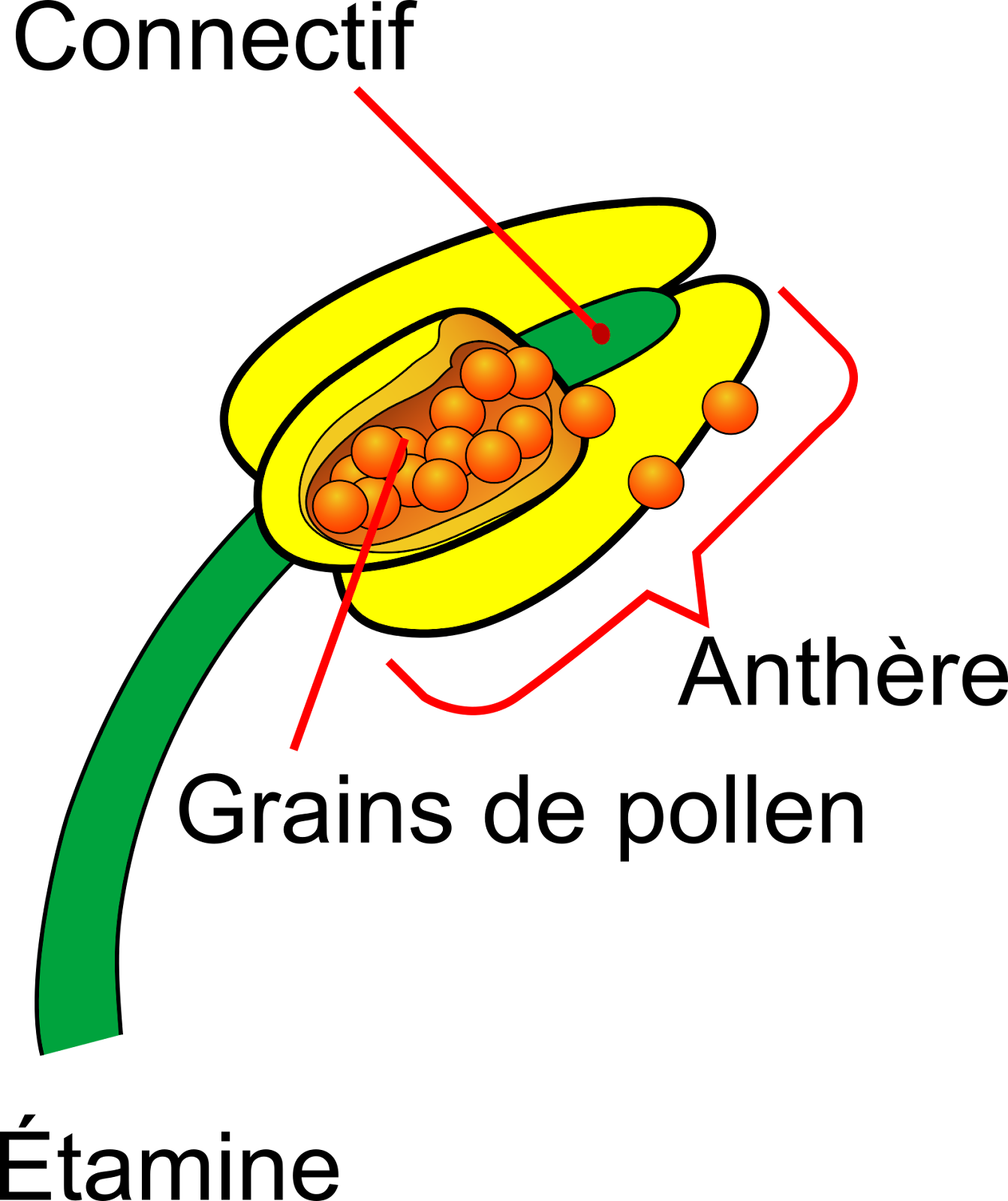
Sur l’avant-dernier verticille se trouve l’appareil reproducteur mâle ou Androcée constitué d’étamines. Une étamine est constituée à sa base d’un filet et à son extrémité de loges renflées appelées des anthères, contenant les grains de pollen qui ne sont autres que des vecteurs de cellules reproductrices mâles. Sur les anthères, on distingue une fente de déhiscence c’est-à-dire une fente d’ouverture qui permet de libérer les grains de pollen. Quand la fente est dirigée vers l’intérieur de la fleur, du côté du pistil, l’étamine est dite introrse. Si la fente est dirigée vers l’extérieur, l’étamine est dite extrorse. Les étamines peuvent être en position alterne par rapport aux pétales et se trouvent alors à la jonction de deux pétales.
Document 8 : Structure d’une étamine
Mature flower diagram-fr.svg, par Mariana Ruiz LadyofHats, translation by Berrucomons ; via Wikimédia Commons, domaine publique, modifié par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Mature_flower_diagram-fr.svg?uselang=fr
Au centre de la fleur, l’appareil reproducteur femelle ou gynécée, appelé aussi pistil, contient les cellules reproductrices femelles, les ovules. Le pistil se divise en trois parties : l’ovaire à la base, le style et l’extrémité appelée le stigmate. Le rôle du stigmate est de récupérer les grains de pollens. Le pistil peut être constitué de un ou plusieurs carpelles soudés ou pas. Un carpelle est une feuille spécialisée dans la production d’ovules et enroulée sur elle-même. La partie basse, l’ovaire, est spécialisé dans la production d’ovules qui suite à l’enroulement apparaîtront en binôme. La partie haute se resserre pour former le style et le stigmate.
Document 9 : Structure du pistil

Mature flower diagram-fr.svg, par Mariana Ruiz LadyofHats, translation by Berrucomons ; via Wikimédia Commons, domaine publique, modifié par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Mature_flower_diagram-fr.svg?uselang=fr
L’ovaire est dit infère quand il est situé dans la partie inférieure de la zone d’insertion des sépales et des pétales. Il sera alors adhérent ou non au réceptacle floral. Il est dit supère quand il est entièrement situé au-dessus de la zone d’insertion des sépales et des pétales.
Document 10 : Différentes positions de l’ovaire dans la fleur : infère à gauche ou supère à droite
Ovary position.svg, par Ulf Mehlig via Wikimédia commons, CC-BY-SA-2.5, modifié par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Ovary_position.svg


Il existe trois sortes de placentation (disposition des ovules dans l’ovaire).
- La placentation axile c’est-à-dire que les placentas sont situés autour d’un axe réunissant les loges de l’ovaire
- la placentation centrale c’est-à-dire que les placentas et les ovules sont en situation centrale et il n’y a plus de paroi entre les carpelles, faisant que l’ovaire ne soit plus constitué que d’une seule loge
- la placentation pariétale c’est-à-dire les placentas et les ovules sont disposés sur la périphérie de l’ovaire en raison du fait que les carpelles ne se soient pas totalement enroulés.
Document 11 : Différentes placentations de gauche à droite : axile, centrale et pariétale



Gynoecium morphology placentation axile-2 cross section.png, par Michael G. Simpson. Redessiné et couleur: Utilisateur: RoRo, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Gynoecium_morphology_placentation_axile-2_cross_section.png
Gynoecium morphology placentation free-central cross section.png, par Michael G. Simpson. Redessiné et couleur: Utilisateur: RoRo, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Gynoecium_morphology_placentation_free-central_cross_section.png
Gynoecium morphology placentation parietal-1 cross section.png, par Michael G. Simpson. Redessiné et couleur: Utilisateur: RoRo, via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Gynoecium_morphology_placentation_parietal-1_cross_section.png
L’étude d’une fleur amène à établir sa formule florale dont l’écriture respecte une codification internationale.
Document 12 : Codification pour la formule florale

On va également disséquer la fleur et produire un diagramme floral
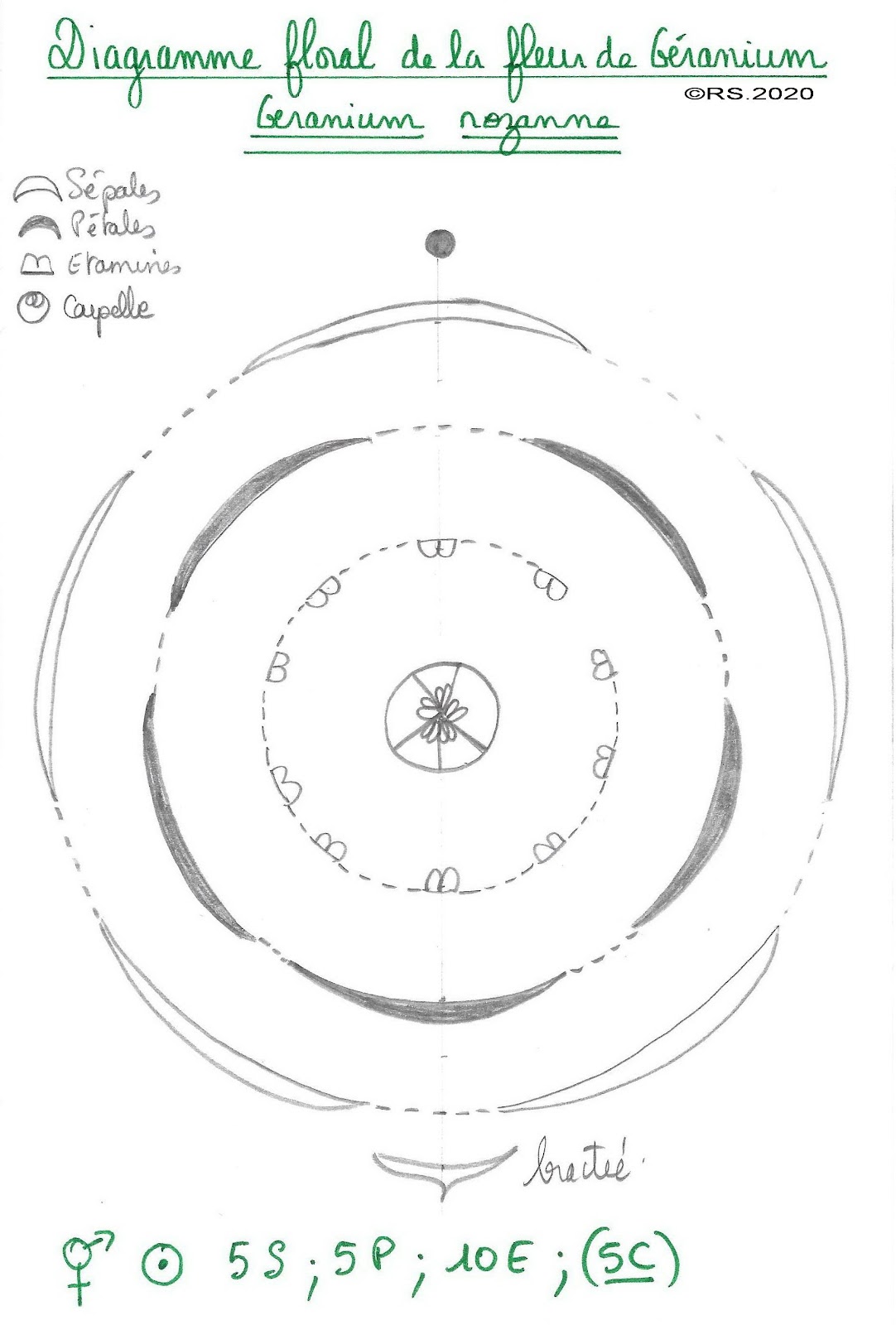
Document 13 : Dissection de la fleur de géranium rozanne

Document 14 : Diagramme floral de la fleur de géranium rozanne
III La fécondation
A/ La rencontre des gamètes
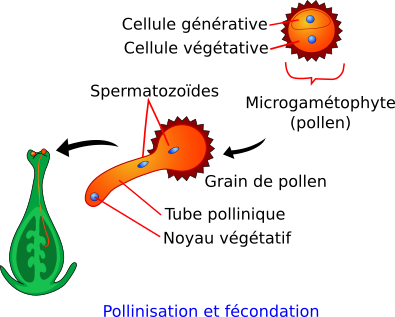
Dans les anthères après la méiose, chaque cellule haploïde s’individualise et sa paroi externe (l’exine) se rigidifie. Cette cellule haploïde subit une mitose asymétrique qui conduit à la formation d’une grande cellule végétative et d’une petite cellule générative. On parle de grain de pollen ou encore de microgamétophyte.
Document 15 : Structure du grain de pollen

Une fois le grain de pollen déposé sur le pistil, la cellule végétative qu’il contient va se déformer et donner un tube pollinique. Ce dernier va creuser le style, passer entre les cellules de celui-ci et descendre jusqu’aux ovaires pour trouver l’ovule au niveau duquel il entrera par un orifice appelé micropyle.
La cellule générative quand à elle, migre dans le cytoplasme de la cellule végétative puis subit une nouvelle mitose pour donner deux cellules spermatiques : les gamètes mâles. Le moment où cette mitose s’effectue dépend des espèces : elle se produit soit avant la dissémination du pollen soit au cours de la croissance du tube pollinique dans le style.
Document 16 : Grains de pollen de campanule déposés sur le stigmate

©RS.2020
Document 17 : Tube pollinique
Mature flower diagram-fr.svg, par Mariana Ruiz LadyofHats, translation by Berrucomons ; via Wikimédia Commons, domaine publique, modifié par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Mature_flower_diagram-fr.svg?uselang=fr
Document 18 : Coupe de pistil traversé par des tubes polliniques

L’ovule des plantes à fleur est plus complexe que chez les animaux. C’est une structure et non une cellule. L’ovule est relié à la paroi ovarienne par le funicule. Il est entouré de 2 téguments qui entourent un tissu nourricier, le nucelle. Les 2 téguments ne se rejoignent pas et l’ouverture laissée se nomme le micropyle. Au sein du nucelle se trouve le sac embryonnaire ou macrogamétophyte.
Document 19 : Structure de l’ovule

Ovule-Gymno-Angio-fr.svg par v via Wikimedia commons, Domaine publique, https://commons.wikimedia.org/wiki/File:Ovule-Gymno-Angio-fr.svg
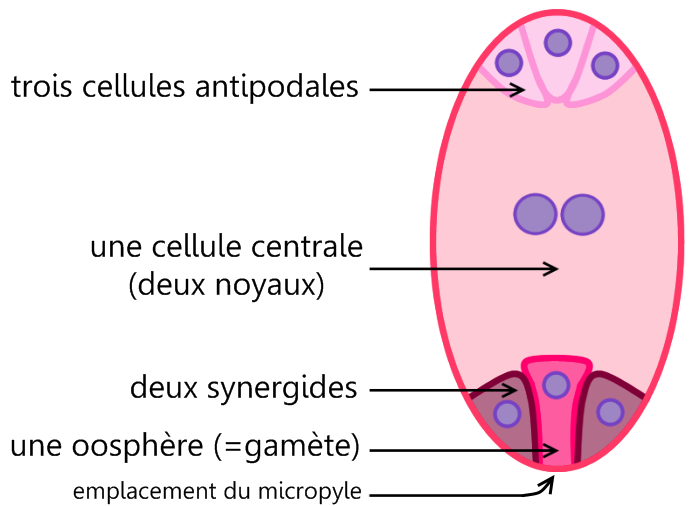
Le sac embryonnaire est constitué de 7 cellules :
- 2 synergides qui encadrent l’oosphère ou gamète femelle au niveau du micropyle
- 3 antipodes au pôle opposé au micropyle
- Une grande cellule centrale qui contient 2 noyaux polaires volumineux.
Document 20 : Structure du sac embryonnaire
Source : Sac-embryonnaire.png par v via Wikimedia commons, CC-BY-SA-3.0-migré, https://commons.wikimedia.org/wiki/File:Sac-embryonnaire.png
Le tube pollinique déverse son contenu dans l’une des synergides et les deux gamètes mâles interviennent l’un et l’autre dans la fécondation du même sac embryonnaire : on parle de double fécondation.
L’un d’eux féconde l’oosphère, l’autre s’unit aux deux noyaux polaires de la cellule centrale du sac embryonnaire. La première fécondation aboutit à la formation d’un zygote diploïde qui évoluera en embryon et la deuxième fécondation donnera un zygote triploïde qui évoluera en albumen, tissu de réserve.
B/ L’autofécondation
L’autofécondation ou autogamie correspond à l’union d’un gamète mâle et d’un gamète femelle d’une même fleur, fleur forcément hermaphrodite. L’autofécondation n’est pas majoritaire chez les angiospermes. Elle est cependant obligatoire pour certaines fleurs qui seront alors qualifiées de cléistogames. On peut citer en exemple le petit pois pour lequel les fleurs ne s’épanouissent jamais entièrement obligeant l’autofécondation : les organes reproducteurs sont enfermés dans une gaine, la carène, formée par deux pétales soudés ;
Document 21 : Fleur de petit pois

©RS.2018
L’autofécondation présente de nombreux avantages.
Tout d’abord, elle permet à l’espèce de se reproduire en absence de pollinisateurs. On peut citer en exemple la fleur de lamier amplexicaule. Cette plante est capable de s’épanouir en plein hiver si le temps est doux et si le sol ne gèle pas. À cette saison les pollinisateurs sont absents et une pollinisation entomogame est donc impossible.
Document 22 : Fleur de Lamier amplexicaule

Henbit Dead-nettle (Lamium amplexicaule) (8526481930).jpg, par Bernard DUPONT from FRANCEvia wikimedia commons, CC-BY-SA-2.0, https://commons.wikimedia.org/wiki/File:Henbit_Dead-nettle_(Lamium_amplexicaule)_(8526481930).jpg?uselang=fr
En pratiquant l’autogamie, la plante assure un fort taux de pollinisation et la quantité de pollen nécessaire est alors moins importante que dans le cas d’une pollinisation entomogame (par les insectes) ou anémogame (par le vent). Il y a donc économie de ressources.
Cette facilité de reproduction permet à l’espèce d’envahir rapidement le biotope assurant ainsi sa survie. En effet, un individu introduit seul dans un nouveau biotope peut se propager en l’absence d’autres plantes de son espèce. L’autopollinisation est donc particulièrement bénéfique pour les plantes peu exigeantes et pouvant se développer dans des milieux pauvres, appelées plantes pionnières. De plus, si l’espèce présente un génotype avantageux, l’absence de brassage génétique dû à l’autopollinisation permet de le conserver.
Document 23 : Lamier amplexicaule envahissant

Lamium amplexicaule3.jpg par KENPEI via Wikimédia Commons, CC-BY-SA-3.0-migré, https://commons.wikimedia.org/wiki/File:Lamium_amplexicaule3.jpg
L’autofécondation présente un inconvénient majeur : elle limite la variabilité génétique des individus. Ainsi on observe une diminution progressive de la vigueur de la plante. Cette diminution de la biodiversité génétique induit une réduction de la capacité d’adaptation de l’espèce aux variations de l’environnement.
C/ La fécondation croisée
La fécondation croisée appelée aussi allogamie correspond à la rencontre de deux gamètes provenant de deux individus différents.
Elle favorise la production d’individus génétiquement nouveaux grâce à des recombinaisons de gènes et les plantes-filles possèdent des caractères différents de ceux des plantes-mères qui peuvent parfois s’avérer être avantageux. La fécondation croisée est utilisée en horticulture afin de procéder à des hybridations pour créer des variétés aux caractères recherchés.
Document 24 : Différentes variétés de pensées

Viola x wittrockiana dsc00949.jpg, par David Monniaux Wikimedia commons, CC-BY-SA-3.0-migré, https://commons.wikimedia.org/wiki/File:Viola_x_wittrockiana_dsc00949.jpg
L’allogamie est dépendante de l’intervention d’éléments pollinisateurs comme le vent, les animaux et parfois l’Homme. Ainsi les plantes allogames réalisent majoritairement leur floraison à la belle saison de manière synchrone à la présence d’insectes pollinisateurs.
Les fleurs hermaphrodites peuvent pratiquer l’allogamie car il existe des mécanismes faisant obstacle à l’autofécondation.
- Mécanismes limitant l’autogamie
La fécondation croisée s’effectue car les fleurs présentent des obstacles à l’autogamie.
On distingue tout d’abord les obstacles morpho-anatomiques qui, comme leur nom l’indique, ont attrait à la structure anatomique des fleurs.
Le premier obstacle correspond à la séparation des organes reproducteurs mâles et femelles dans l’espace. Ainsi une plante, en portant à la fois des fleurs unisexuées mâles et des fleurs unisexuées femelles, diminuera la probabilité d’autofécondation. L’espèce est alors dite monoïque. Cela concerne 7 % des angiospermes et on peut citer en exemple des plantes dans la famille des Fagacées, le hêtre et le chêne, dans la famille des Bétulacées, le bouleau ou encore quelques espèces de Graminées comme le maïs.
Dans 4 % des genres, les fleurs unisexuées sont portées par des individus différents. L’espèce est alors dite dioïque. On peut citer en exemple l’ortie, le peuplier et l’asperge. La dioécie, c’est-à-dire le fait que l’espèce soit dioïque, rend de fait l’autogamie impossible.
Document 25 : Répartition des sexes chez les angiospermes
Chez certaines espèces hermaphrodites, il existe des barrières mécaniques empêchant l’autofécondation. C’est le cas de la famille des Orchidacées dont les fleurs présentent une fusion partielle de l’androcée et du gynécée à l’origine d’une membrane appelée rostellum qui empêche les grains de pollen des anthères appelée pollinies, de rencontrer les stigmates.
Document 26 : Fleur d’orchidée

À ce propos, on peut raconter l’histoire de la vanille Bourbon qui a été introduite en 1919 à l’île de la Réunion dans un but agricole. L’espèce en provenance du Mexique, pourtant bien acclimatée n’a jamais pu produire de gousses de vanille au grand désespoir des colons installés. La raison était qu’aucun insecte sur l’île n’arrivait à la polliniser. C’est un jeune esclave de 12 ans Edmond Albius qui découvrit comment polliniser la plante en l’absence de son pollinisateur naturel. Il suffit à l’aide d’une petite baguette de bois, d’écarter le rostellum et de presser la fleur pour mettre en contact les parties mâles et femelles.
Il existe également des obstacles physiologiques à l’autofécondation. C’est le cas par exemple, quand la maturité des organes reproducteurs est décalée dans le temps. Les étamines sont mûres soit avant le pistil, on parle alors de protandrie, soit après le pistil et on parle alors de protogynie. La protandrie est la modalité la plus fréquente.
C’est le cas par exemple chez la campanule à grandes fleurs. Les étamines introrses sont au début de leur maturation accolées au pistil. À maturité, leur fente de déhiscence laisse s’échapper du pollen qui se déposera alors sur le style poilu du pistil. Au fur et à mesure de la maturation des étamines, celles-ci se courbent, éloignant les anthères du pistil encore non mature. Quand les ovules sont prêts, les cinq stigmates activés de cette fleur se courbent, présentant ainsi une extrémité gluante permettant la fixation du pollen. Les anthères étant vides il y a peu de probabilités d’autopollinisation. Le décalage de maturité dans le temps évite donc l’autofécondation. Cependant si aucun des stigmates n’a été pollinisé, ces derniers poursuivent leur courbure jusqu’à toucher le style et récupérer ainsi du pollen déposé par les anthères au moment de leur maturation. Par ce procédé d’autofécondation tardif, la plante assure la survie de l’espèce.
Document 27 : Fleurs de campanules

On peut citer enfin les obstacles génétiques comme la stérilité mâle de la fleur qui correspond à une incapacité de la plante à produire du pollen fécondant. Ce mécanisme est cependant rare et ce sont plutôt des mécanismes d’auto-incompatibilité pollen/pistil qui sont observés. Ces mécanismes se situent au niveau de la germination ou du développement du grain de pollen sur les stigmates. Ces systèmes font appel à des gènes d’incompatibilité (S) existant sous forme de nombreux allèle (S1, S2, S3,…).
Il existe une auto-incompatibilité gamétophytique c'est-à-dire liée aux gamètes. Les plantes hétérozygotes diploïdes disposent pour le gène S de deux allèles. Le gamétophyte mâle haploïde ne contient qu’un seul de ses allèles. Si l’allèle du pollen est le même que l’un des deux allèles de l’ovaire, le développement du tube pollinique sera bloqué et la fécondation ne pourra avoir lieu. Ainsi seule la fécondation entre une plante et un grain de pollen disposant d’allèles différents est possible.
Il existe une auto-incompatibilité sporophytique c'est-à-dire liée à la plante. Elle s’observe dans le cas où une partie des composants de la paroi du pollen a été synthétisée par les cellules nourricières des anthères au cours de sa formation. Dans ce cas les cellules entrant en contact avec les stigmates sont des cellules diploïdes possédant alors deux allèles. Si l’un des deux allèles de la plante diploïde qui a généré le pollen est le même que l’un des deux allèles de l’ovaire de la plante receveuse, alors le développement du tube pollinique sera bloqué et la fécondation ne pourra avoir lieu. Seule la fécondation entre deux plantes éloignées disposant d’allèles différents est possible.
Document 28 : Auto-incompatibilité gamétophytique (GSI) et sporophytique (SSI)
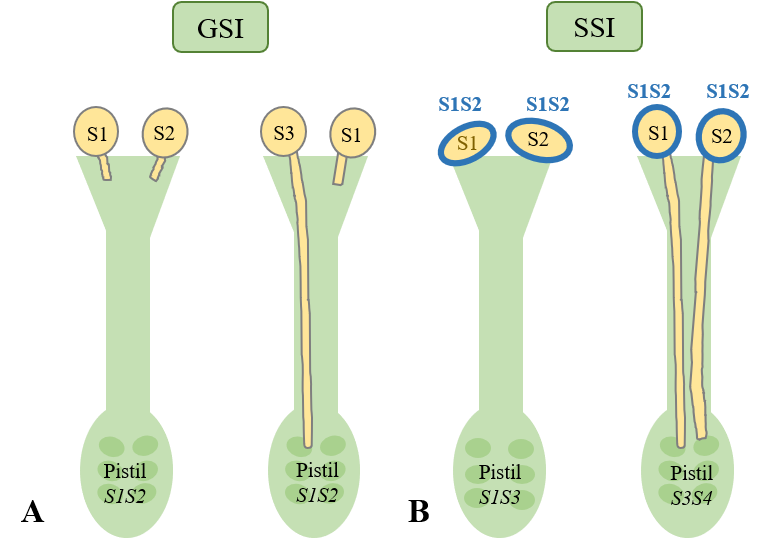
Auto-incompatibilité pollinique.png par Primroses32 via Wikimédia Commons, CC-BY-SA-4.0
https://commons.wikimedia.org/wiki/File:Auto-incompatibilit%C3%A9_pollinique.png
- La pollinisation
La pollinisation correspond au transport du pollen vers les organes reproducteurs femelles.
Il existe différents types de vecteurs. Les fleurs vont donc être qualifiées selon leur mode de pollinisation.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Les fleurs anémogames sont des fleurs discrètes souvent verdâtres et inodores. Chez les Graminées, elles sont regroupées en épi et présentent deux stigmates plumeux et des étamines flexueuses exposées à la surface de l’épi. Chez les arbres forestiers les fleurs mâles sont regroupées en inflorescences appelées « chatons ».
Document 29 : Fleur de graminée

Grassflower-fr.svg, Original:David CondreyVecteur: Spedona, via Wikimedia commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Grassflower-fr.svg
Document 30 : Épi de graminée présentant des étamines pendantes

Briza media spikelet.jpg, par v via wikimedia commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Briza_media_spikelet.jpg
La déhiscence des anthères libèrent de grandes quantités de pollen lisse et de très petite taille (10 à 15 nm) qui peut être transporté par le vent sur de très grandes distances. Les pertes en grains de pollen sont énormes mais la quantité libérée compense le côté aléatoire de la rencontre pollen – stigmates. Les stigmates plumeux ont pour rôle, tel un peigne, de capter les grains de pollen.
Document 31 : Pollen de graminée, MO x400
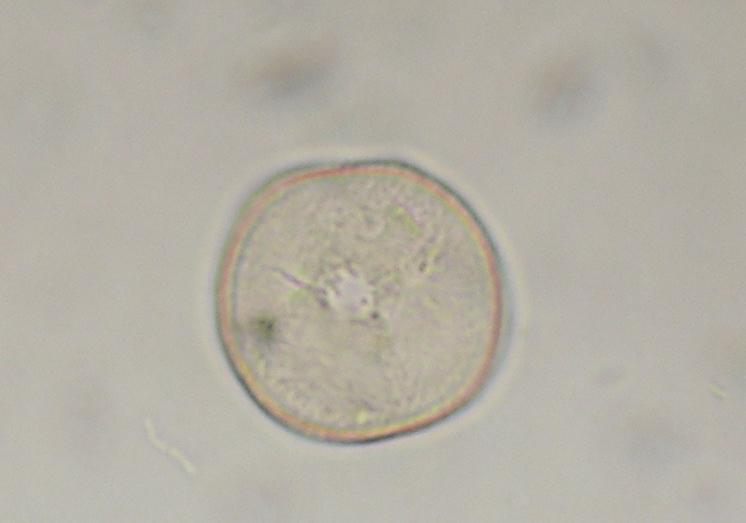
Pollen d'herbe (6864642413) .jpg, par v via Wikimedia commons, CC-BY-SA-2.0, https://commons.wikimedia.org/wiki/File:Grass_Pollen_(6864642413).jpg
Les fleurs zoogames dont le pollen gros et collant est transporté par les animaux, présentent en général des fleurs odorantes aux pétales colorés et possédant des nectaires.
Document 32 : Grains de pollen d’hibiscus dont l’exine présente une ornementation permettant de se fixer aux insectes pollinisateurs
©RS.2020

Les fleurs entomogames sont très fréquentes et représentent 90 % des espèces chez les angiospermes. Elles possèdent généralement des pétales colorés en jaune, orange (caroténoïdes), en bleu, rouge, pourpre ou rose (anthocyanes), des couleurs visibles par les oiseaux et les insectes dont le spectre de sensibilité visuelle va de l’ultraviolet à l’orange.
Document 33 : Variété de couleurs des pétales chez les angiospermes

Affiche fleur 2.jpg, par Alvesgaspar, via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Flower_poster_2.jpg
Document 34 : Spectre de sensibilité visuelle des pinsons
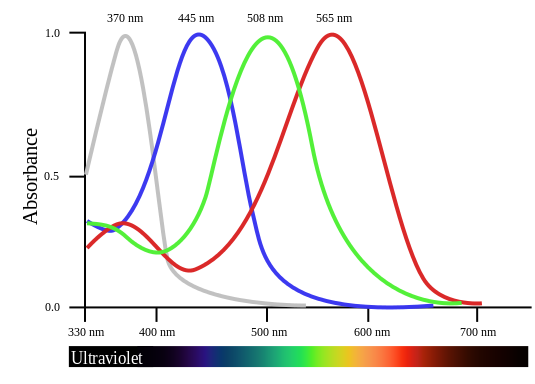
BirdVisualPigmentSensitivity.svg, par L. Shyamal via Wikimedia commons, domaine publique, https://commons.wikimedia.org/wiki/File:BirdVisualPigmentSensitivity.svg
Les insectes ont une sensibilité très développée pour les composés volatiles émis par les fleurs : les terpènes. On peut citer le géraniol du rosier, le limonène du citronnier ou encore certains composés comme la vanilline des orchidées.
Document 35 : Géraniol du rosier à gauche et limonène du citronnier à droite

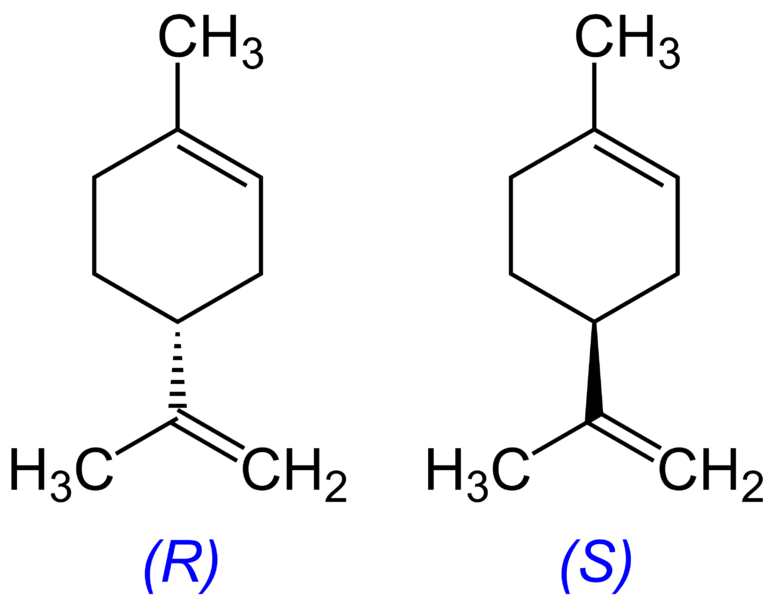
Geraniol.png par Jan Herold , Leyo via Wikimedia commons, domaine publique, https://commons.wikimedia.org/wiki/File:Geraniol.png
Le nectar produit à partir des nectaires est une ressource alimentaire pour les insectes. Formé à partir du phloème, il est constitué d’eau, de sucre, d’acides aminés, et attire les pollinisateurs.
Enfin il a été observé des mécanismes résultant d’une coévolution. Dans le cas de la sauge, les étamines s’abaissent quand l’abeille s’introduit dans la fleur pour boire le nectar produit au fond de la corolle tubulaire. Ce mécanisme de bascule permet de déposer du pollen sur le dos de l’insecte afin de faciliter la dispersion de celui-ci.
Document 36 : Fleur de sauge et ses 2 étamines basculantes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
sage-1598405_1920, par Image par kie-ker de Pixabay , Pixabay licence, libre pour usage commercial, https://pixabay.com/fr/photos/sage-fleur-sauvage-sauge-salvia-1598405/
Fleurs et pollinisation- SVT - ENJEUX Term spé #7 - Mathrix
Date de dernière mise à jour : 10/05/2022