La production de matière par les plantes à fleurs
Vidéo en bas de page ^^
I Rappels et contexte d’étude :
Une plante à fleur est constituée d’un appareil végétatif comprenant des racines ancrées dans le sol et des tiges feuillées se développant en milieu aérien.
Document 1 : Organisation générale d’une plante à fleurs
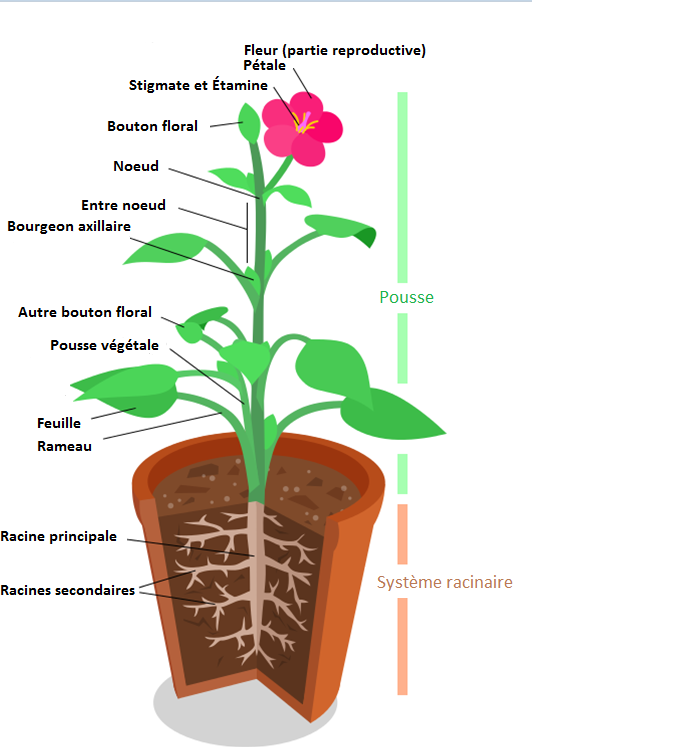
Source : Schéma plante + fleur.png, par Fatlord , via Wikimedia commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Sch%C3%A9ma_plante_%2B_fleur.png
Les plantes à fleurs ont une vie fixée à l’interface sol/atmosphère pour les plantes aériennes ou sol/eau pour les plantes aquatiques. On les trouve dans des environnements variables et doivent donc résister aux variations du milieu.
Les plantes à fleurs sont des végétaux chlorophylliens et n’ont pas besoin de matière organique pour vivre car elles réalisent la photosynthèse, qui est la production de matière organique (amidon) dans les parties aériennes de la plante, en présence de lumière, à partir d’eau et du dioxyde de carbone de l’air.
Un système de vaisseaux conducteurs permet le transport de l’eau et des sels minéraux (sève brute) des racines vers les parties aériennes et de la matière organique et de l’eau (sève élaborée), des parties chlorophylliennes vers le reste de la plante
II. Localisation de la production de matière organique
La photosynthèse est la fabrication (la synthèse) de sucre à partir de matière minérale grâce à l’énergie lumineuse. Elle permet donc leur entrée dans l’ensemble du monde vivant (biosphère). La photosynthèse a principalement lieu dans les feuilles qui forment une grande surface d’échange avec l’atmosphère ou avec l’eau. Elles y prélèvent le dioxyde de carbone et captent la lumière grâce aux chloroplastes, rejettent de la vapeur d’eau et du dioxygène.
Equation bilan de la photosynthèse :
6 CO2 +6 H2O C6H12O6 + 6O2
Document 2 : Molécule de glucose
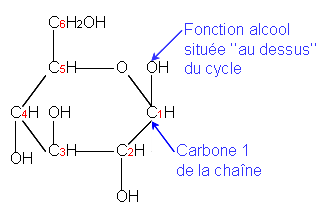
Source : Béta-glucose.PNG, par Belgarath007, CC-BY-SA-3.0-migrated, via Wikimédia Commons, https://commons.wikimedia.org/wiki/File:B%C3%A9ta-glucose.PNG?uselang=fr
La matière organique produite est donc du sucre précisément du glucose. Or quand on mange des feuilles comme des épinards ou de la salade on ne ressent pas le goût sucré. Cela signifie donc que le glucose produit est soit consommé de suite par la plante soit stocké sous la forme d’un sucre n’activant par les récepteurs du goût de notre langue à savoir un sucre complexe et probablement de l’amidon.
L’amidon est facilement mis en évidence par un réactif le Lugol appelé aussi eau iodée. Celui-ci de couleur brun jaunâtre devient noir en sa présence.
Pour vérifier cette hypothèse de stockage sous forme d’amidon une expérience très simple peut être réalisée. Prenons deux plants d’élodée, une plante à fleurs aquatique, plaçons-en une à l’obscurité et l’autre à la lumière pendant trois jours. Récupérons les feuilles et versons de l’eau iodée dessus puis observons le résultat au microscope. On peut constater que la plante placée à l’obscurité ne présente pas d’amidon alors que celle placée à la lumière en possède et qu’il est précisément localisé dans les chloroplastes. Ainsi si la photosynthèse productrice de glucose a lieu dans le chloroplaste c’est également dans cet organite que ces molécules de sucre simple (C6H12O6) s’associent entre elles pour former de l’amidon (C6H10O5)n.
Document 3 : Molécule d’amidon (amylopectine)

Source : Structure de l'amylopectine.JPG par Laranounette via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Structure_de_l%27amylopectine.JPG
Le chloroplaste est un organite délimité par une double membrane. Son stroma, milieu intérieur, présente un réseau de membranes formant des sacs aplatis appelés thylakoïdes. Chaque thylakoïde délimite un espace intra –thylakoïdien. Ces thylakoïdes s’empilent pour former ce qu’on appelle un granum. Nous verrons plus tard quel est le rôle de ces structures particulières. Dans le stroma d’un chloroplaste, on distingue donc plusieurs grana, des grains d’amidon mais également des gouttelettes lipidiques.
Document 4 : Chloroplaste observé au microscope électronique
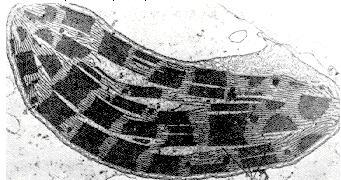
Source : Plast.JPG, Wikimédia Commons, licence public GNU, https://commons.wikimedia.org/wiki/File:Plast.JPG
Document 5 : Structure d’un chloroplaste

Légende :
(1) membrane externe ; (2) espace intermembranaire ; (3) membrane interne ; (4) stroma ; (5) lumen du thylakoïde ; (6) membrane du thylakoïde ; (7) granum (empilement de thylakoïdes) ; (8) thylakoïde ; (9) amidon ; (10) ribosome ; (11) ADN chloroplastique ; (12) plastoglobule (gouttelette lipidique).
Source : Par SuperManu — own work based on Chloroplaste-schema.gif, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=2923179
Document 6 : Coupe de Chlamydomonas au microscope électronique transmission révélant des chloroplastes.

Source : Chlamydomonas TEM 07.jpg, domaine publique via Wikimedia commons, https://commons.wikimedia.org/wiki/File:Chlamydomonas_TEM_07.jpg
III. La matière organique produite lors de la photosynthèse
Les chloroplastes sont donc le lieu de la production de matière organique : de l’amidon est produit dans le stroma. Existe-t-il d’autres produits de la photosynthèse et si oui quels sont-ils ?
Entre 1950 et 1960, trois chercheurs, Calvin, Benson et Bassham réalisèrent une série d’expériences qui leurs permirent d’élucider les étapes chimiques qui conduisent, à partir du dioxyde de carbone, aux premières molécules organiques lors de la photosynthèse. Ils travaillèrent sur des chlorelles. Ces algues unicellulaires ont été mises en culture dans un milieu enrichi en dioxyde de carbone et éclairées afin de faciliter la photosynthèse. Ils eurent l’idée d’utiliser du carbone radioactif pour tracer les molécules produites lors de la photosynthèse. En effet, ce carbone radioactif étant utilisé par les cellules chlorophylliennes pour réaliser la production de matière, les molécules carbonées produites seront repérables par radioactivité.
Document 7 : Expérience de Calvin, Benson et Bassham

Pour pouvoir identifier et nommer les molécules produites uniquement par la photosynthèse et surtout pour les distinguer des autres, il faut avant tout faire la liste des molécules présentes dans les cellules. Pour cela, il suffit de laisser les cellules incuber le plus longtemps possible en présence dioxyde de carbone normal et de lumière. Une fois la suspension de Chlorelles fixée par l’éthanol bouillant (les cellules éclatent mais les composés qu’elles contiennent ne sont pas détruits), il suffit de réaliser une séparation des constituants par chromatographie bidimensionnelle à l’aide de 2 solvants différents.
Après avoir réalisé le dépôt de suspension de chlorelles fixées à l’alcool sur le support de chromatographie, une première séparation est effectuée à l’aide d’un solvant constitué d’eau et de phénol. Les molécules remontent le papier selon leur taille et leur masse. Ainsi apparaissent plusieurs tâches. Cependant chacune d’entre elles peut contenir des composés de taille et de masse identiques et il conviendra alors de les séparer. Pour cela il suffit de pivoter le papier à 90°, de le plonger dans un autre solvant constitué de butanol et d’acide propionique afin de les séparer. Toutes les molécules seront ainsi séparées dans l’espace.
La révélation du chromatogramme se fera par différents colorants spécifiques (pour les curieux : acides organiques par le bleu de bromophénol, acides aminés par la ninhydrine, sucres par le naphtorésorcinol et composés phosphorylés par le molybdate d'ammonium). Ainsi nous obtenons une chromatographie bidimensionnelle servant de témoin et permettant d’identifier toutes les substances solubles présentes dans les chlorelles.
Document 8 : Schématisation du résultat de la chromatographie bidimensionnelle des substances contenues dans les chlorelles et révélées par des colorants spécifiques.
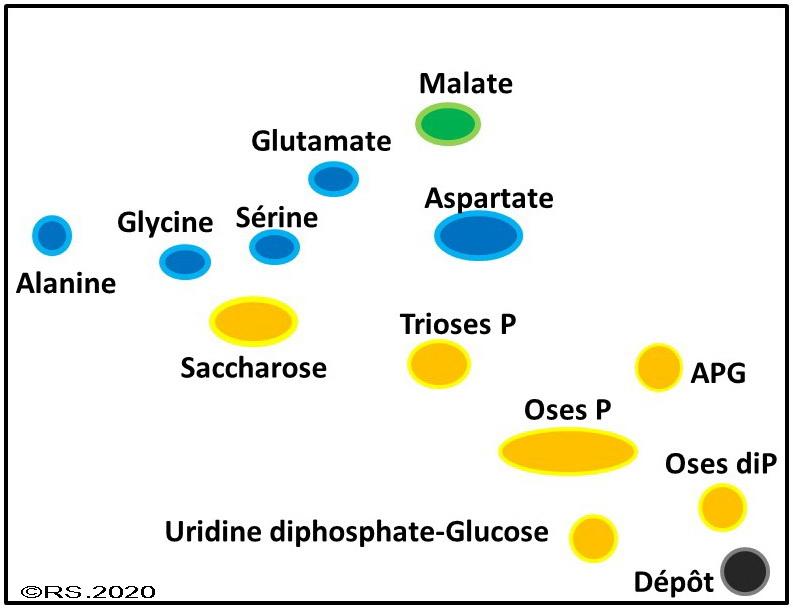
L’objectif est donc d’identifier parmi toutes ces substances, lesquelles ont été produites par la photosynthèse et si possible dans quel ordre. Pour ce faire, l’expérimentation a été réalisée plusieurs fois de suite en arrêtant la photosynthèse à des temps de plus en plus longs après l’injection du carbone radioactif. Sur le dispositif expérimental, la durée d’exposition au carbone 14 dépendra du lieu d’injection de celui-ci sur le trajet amenant les chlorelles à l’alcool bouillant. Si vous l’injectez au point le plus bas du circuit, les chlorelles auront eu peu de temps pour l’utiliser dans leur processus photosynthétique avant d’être éclatées. Si vous l’injectez au point le plus haut du circuit, les chlorelles auront eu beaucoup de temps pour l’utiliser.
C’est comme quand vous jouez au p’tit bac ! Vous savez ? Ce jeu où il faut trouver des mots de différentes catégories commençant par une lettre précise ? Pour savoir quelle lettre sera sélectionnée, vous demandez à un joueur de réciter l’alphabet dans sa tête et vous dites stop ! Si vous dites stop rapidement vous tomberez sur une lettre située en début d’alphabet si vous attendez plus longtemps vous serez plus avancés dans l’alphabet.
Et bien ici c’est pareil. Si vous stoppez rapidement les chlorelles après l’injection de carbone radioactif, elles seront peu avancées dans les réactions de la photosynthèse. Alors, que si vous attendez plus longtemps, le processus sera plus avancé et vous verrez des molécules différentes de celles du début. En stoppant méthodiquement la photosynthèse, vous pourrez ainsi reconstituer les étapes de la production de matière.
Les réactions ont ainsi été stoppées à 2, 5, 10 et 30 secondes. Les chromatographies ont été réalisées. Si on les colorait comme précédemment, on ne pourrait pas être sûr que toutes les substances révélées seraient celles provenant de la photosynthèse. Le seul moyen de révéler les produits de la photosynthèse, est de positionner les chromatogrammes contre un film photographique à l’obscurité. Les rayons émis par le carbone 14 intégré dans les molécules produites, vont impressionner la plaque photographique et une tâche apparaîtra. Chaque tâche indique donc la présence d’un composé radioactif formé par photosynthèse à partir d’un carbone radioactif. La comparaison des chromatogrammes colorés et des chromatogrammes révélés par autoradiographie permet donc de savoir quels produits ont été synthétisés au cours du contact avec le carbone 14.
Document 9 : Principe de révélation par autoradiographie de la chromatographie bidimensionnelle de Calvin

Document 10 : Résultats obtenus lors de l’expérience de Calvin, Benson et Bassham
Les documents ci-dessous sont des schématisations des autoradiographies obtenus par les 3 scientifiques.
On constate qu’après moins de 2 secondes la première molécule formée est un composé en trois carbones appelé l'APG ou acide PhosphoGlycérique.




À 5 secondes on constate qu’il y a eu production de pentoses (sucre à 5 carbones) : le Ribulose-phosphate RUP (identifié Oses P) et le Ribulose-biphosphate ou RUBP (identifié Oses diP).
À 10 puis à 15 secondes, on observe en plus l’apparition de triose-phosphate, de malate et d’acides aminés (glycine, alanine….)
L'acide phosphoglycérique (APG) est donc le premier composé formé : il est à l'origine de toutes les synthèses.
Le produit de la photosynthèse c’est le glucose, un sucre à six carbones. Il est produit à partir du dioxyde de carbone et on pourrait donc s’attendre à ce qu’un dioxyde de carbone soit fixé sur une molécule à cinq carbones pour en faire une à 6 carbones. Or un pentose (C5) bisphospate (le RUBP ou Ribulose bis-phosphate) peu connu apparaît rapidement dans les radiochromatogrammes. C'est donc lui l'accepteur de CO2. Ainsi le chloroplaste disposerait d’une réserve de sucre à cinq carbones qui joueraient le rôle d’accepteur de dioxyde de carbone à l’origine de la production d’une première molécule l’APG. Or l'acide phosphoglycérique (APG) n'est pas un sucre. La tâche formée par ce composé reste quasi identique quel que soit le temps, contrairement à la taille des tâches des trioses-phosphates et des sucres en C5 qui ne fait qu’augmenter traduisant une accumulation dans les microalgues de ces catégories de sucres. Comme de nouveaux produits sont apparus mais que la tâche d’APG reste constante sur les différents chromatogrammes, on peut donc penser qu’il n’est pas exporté pour la consommation cellulaire mais qu’il interviendrait comme molécule intermédiaire dans le processus de fabrication. Pour entrer dans les réactions de synthèse des composés organiques, l’APG doit être réduit en triose-phosphate (……) qui apparait effectivement rapidement dans les radiochromatogrammes.
Pour que l'ensemble fonctionne, il faut que le RUBP (qui existe en faible quantité) soit régénéré et ceci ne peut se faire qu'à partir des composés dérivés des trioses-phosphate.
L'incorporation du CO2 se réalise donc globalement de la manière suivante :
RUBP + CO2 2APG
2 Trioses phosphate
RUBP
Incorporation réduction régénération
D’autres expériences ont mis en évidence la consommation de 18 molécules énergétiques, l’ATP (adénosine triphosphate) ainsi que de 12 molécules fournisseuses d’ions H+, le NADPH2.
Le nicotinamide adénine dinucléotide phosphate (NADP) est un coenzyme présent dans toutes les cellules vivantes. Il s'agit d'un dinucléotide, car il est constitué d'un premier nucléotide, dont la base nucléique est l'adénine, uni à un second nucléotide, dont la base est le nicotinamide. Le NADP existe sous une forme réduite, notée NADPH2, et une forme oxydée, notée NADP+. Le NADP intervient dans le métabolisme comme transporteur d'électrons dans les réactions d'oxydoréduction, le NADPH2 comme réducteur et le NADP+ comme oxydant. On a donc affaire ici à une réaction d’oxydo-réduction.
Un cycle apparaît alors.
Dans ce système il faut produire un sucre à 6 carbones. Or la dernière étape du cycle passe de 2 sucres à 3 carbones (soit 6 au total) à une molécule à 5 carbones (RUBP). Un carbone est rendu disponible. Ainsi en répétant 6 fois ce cycle, on obtient 12 trioses-phosphates de libres et on peut produire un hexose (sucre à 6 carbones) visible à 30 secondes : l’Uridine diphosphate-Glucose (UDPG).
Le sucre produit est soit consommé pour les réactions métaboliques du cytoplasme, soit stocké sous forme d’amidon, soit utilisé pour former du saccharose, forme de transport des sucres par la sève.
Document 11 : Cycle de Calvin simplifié
Source : Calvin-Zyklus de.svg, par Yikrazuul via wikimédia commons, domaine publique, modifié par Sandra Rivière, https://commons.wikimedia.org/wiki/File:Calvin-Zyklus_de.svg

Pour les curieux, voici en détail le cycle de Calvin :
Étape 1: Fixation du dioxyde de carbone ou Carboxylation par action d’une enzyme, la rubisco.
6 RUBP + 6 CO2 + 6 H2O 12 APG + 12H+
Étape 2 : Réduction de l’APG en triose-phosphate : l’aldéhyde phosphoglycérique
Elle se passe en deux temps :
- phosphorylation de l’acide phosphoglycérique (APG)
Cette phosphorylation nécessite un apport de phosphate inorganique que l’on peut trouver dans les molécules d’ATP ou adénosine triphosphate. L’ATP va perdre un phosphate inorganique Pi et donner de l’ADP adénosine diphosphate. Le phosphate inorganique va se fixer sur l’APG et donner de l’acide biphosphoglycérique.
12 APG + 12 ATP 12 ABPG (acide biphosphoglycérique) + 12 ADP
- réduction de l’acide biphosphoglycérique ABPG
Cette réduction nécessite une molécule réduite fournisseuse d’ions H+. C’est le NADPH. Le nicotinamide adénine dinucléotide phosphate (NADP) est un coenzyme présent dans toutes les cellules vivantes. Il s'agit d'un dinucléotide, car il est constitué d'un premier nucléotide, dont la base nucléique est l'adénine, uni à un second nucléotide, dont la base est le nicotinamide. Le NADP existe sous une forme réduite, notée NADPH2, et une forme oxydée, notée NADP+. Le NADP intervient dans le métabolisme comme transporteur d'électrons dans les réactions d'oxydoréduction, le NADPH2 comme réducteur et le NADP+ comme oxydant. On a donc affaire ici à une réaction d’oxydo-réduction.
12 ABPG + 12 (NADPH + H+) 12 aldéhyde phosphoglycérique (triose-phosphate) + 12 NADP+ + 12 Pi
Étape 3 : Régénération du Ribulose-Biphosphate RUBP
- Recyclage du glycéraldéhyde 3P en ribulose-phosphate
Cette étape consiste à reconvertir les molécules à 3 atomes de carbone en molécules à 5 atomes de carbone afin de pouvoir les réutiliser dans le cycle. L’équation bilan est complexe et non étudiée ici. Elle libère 2 phosphates inorganiques.
10 aldéhyde phosphoglycérique (triose-phosphate) + 4 H2O 6 ribulose-phosphate (RuP) + 4Pi
- Régénération du ribulose-biphosphate RUBP
Le RuP a besoin d’un phosphate inorganique supplémentaire et c’est l’ATP qui va l’apporter.
6 RuP + 6 ATP 6 RUBP (Ribulose BiPhostate) + 6 ADP + 6H+
Cette réaction de phosphorylation se fait sous l’action d’une enzyme : la phosphate ribulose kinase.
Une fois le tour de cycle terminé, il reste 2 molécules d’aldéhyde phosphoglycérique (aldPG) non utilisées. Elles serviront pour la production de glucose.
2 aldéhyde phosphoglycérique (triose-phosphate) + H2O Glucose-phosphate
La somme totale des réactions du cycle de Calvin est :
6 CO2 + 12 NADPH2 + 18 ATP + 11 H2O → C6H11O9P2- (glucose-6-phosphate) + 12 NADP+ + 18 ADP + 16 Pi + 6 H+(aq)
avec
Pi = HPO42-
IV. Les mécanismes de la photosynthèse
Calvin a donc mis en évidence que la production de glucose nécessitait l’intervention des molécules énergétiques type ATP ainsi que des molécules appelées coenzymes (NADPH2).
On peut se demander quelle est l’origine de ces deux types de molécules.
Arnon a mis au point une expérience qui a permis de déterminer de quelle partie du chloroplaste provenait ces molécules. Pour cela il a déstructuré des chloroplastes en deux fractions : le stroma et les thylakoïdes.
Il a testé chacune d’elle.
Document 12 : Expérience de Arnon
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
L’expérience 1 lui apprend que le stroma seul ne permet pas la production des molécules organiques par incorporation de CO2.
L’expérience 2 est en quelque sorte une reconstitution d’un chloroplaste hybride dont le stroma serait resté à l’obscurité mais dont les thylakoïdes auraient été éclairés. On sait que la lumière est la source d’énergie des cellules autotrophes. Ainsi si les thylakoïdes sont responsables de la production des molécules énergétiques d’ATP, comme ils ont été éclairés, alors du dioxyde de carbone devrait être fixé. Si c’est le stroma qui est responsable de leur production alors, dans cette expérience, comme le stroma était l’obscurité, il ne devrait pas y avoir d’ATP et donc pas d’incorporation du dioxyde de carbone. On constate qu’il y a fixation de dioxyde de carbone. On peut donc en déduire que les molécules d’ATP sont produites par les thylakoïdes car ce sont les seuls éléments qui ont reçu de la lumière.
L’expérience 3 montre que les thylakoïdes exposés à la lumière peuvent être remplacés par un apport de NADPH2 et d’ATP. On peut donc déduire de cette expérience que les thylakoïdes exposés à la lumière sont bien responsables de la production d’ATP mais aussi de NADPH2, nécessaires au fonctionnement du cycle de Calvin qui a lieu pour sa part dans le stroma du chloroplaste.
Reprenons l’équation bilan globale de la photosynthèse.
Il a été démontré que le glucose est formé grâce cycle de Calvin à partir du CO2. On peut se demander d’où provient le dioxygène libéré. Provient-il à du dioxyde de carbone ou de l’eau ?
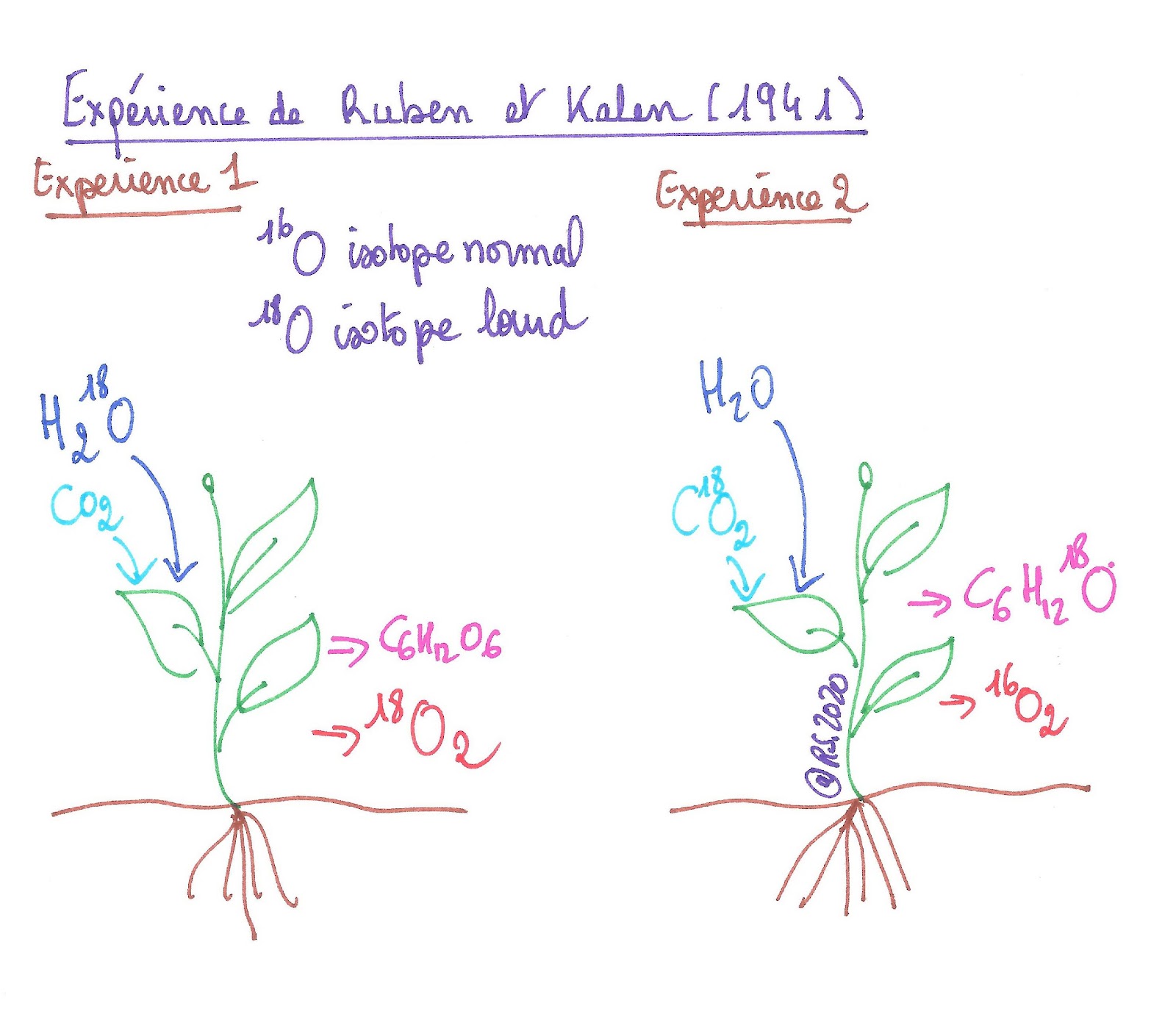
Ruben et Kalen en 1941 ont utilisé un isotope lourd de l’oxygène, le 18O à la place de 16O pour marquer le dioxyde de carbone et l’eau impliquée dans la réaction globale de la photosynthèse. Cela permettait d’identifier de dioxygène rejeté.
En effet en fournissant de l’eau lourde aux feuilles, s’ils observaient du dioxygène lourd à la fin de la photosynthèse alors cela signifiait que c’était l’eau qui avait fourni le dioxygène. S’ils n’observaient pas de dioxygène lourd alors cela signifiait que ce n’était pas l’eau qui fournissait le dioxygène.
Ils ont raisonné de même pour le dioxyde de carbone lourd. Si le dioxygène libéré n’était pas lourd à la fin de la photosynthèse c’est que ce n’était pas le dioxyde de carbone qui le fournissait. Au contraire si le dioxygène libéré à la fin de la photosynthèse était lourd c’est qu’il provenait du dioxyde de carbone.
Document 13 : Expérience de Ruben et Kalen (1941)
Les résultats de leurs expériences ont montré que quand ils fournissaient du dioxyde de carbone lourd aux feuilles, ils obtenaient du glucose lourd et que quand ils fournissaient de l’eau lourde aux feuilles, ils obtenaient du dioxygène lourd. Cette expérience a donc confirmé que le dioxyde de carbone servait à produire le glucose et que l’eau était à l’origine de la libération de dioxygène.
Document 14 : La molécule d’eau
Source : La molécule d'eau.PNG par H'arnet, via Wikimedia Commons, CC-BY-SA-3.0,2.5,2.0,1.0, https://commons.wikimedia.org/wiki/File:La_mol%C3%A9cule_d%27eau.PNG?uselang=fr
Cela signifie donc que les molécules d’eau impliquées dans la photosynthèse ont été brisées. On sait que la photosynthèse ne se réalise qu’en présence de lumière et que briser des liaisons covalentes nécessite de l’énergie. Or la lumière, c’est de l’énergie. On suppose donc il y a eu un mécanisme de lyse de l’eau par la lumière : on parle de photolyse.
C’est Robert Hill, biochimiste anglais, qui en 1930 émit l’hypothèse que la production de dioxygène nécessitait la présence d’un accepteur d’électrons en plus de la lumière car quand on casse une molécule d’eau, un électron est libéré. Il y a donc bien forcément une molécule pour le récupérer.
Réaction d’oxydation de l’eau :
2 H2O O2 + 4H+ + 4e-
On sait que :
2 (NADPH + H+) 2NADP + 4H+ + 4e-
Couplons les réactions :
2 H2O O2 + 4H+ + 4e-
et
2NADP + 4H+ + 4e- 2 (NADPH + H+)
On obtient :
2 H2O + 2NADP O2 + 2 (NADPH + H+)
Or cette réaction n’est pas spontanée et exige de l’énergie qui proviendrait donc de la lumière. Pour vérifier cela, il étudie le dégagement de dioxygène d’une suspension de chloroplaste en présence et en absence de lumière et d’un réactif portant son nom, le réactif de Hill, du potassium hexacyanoferrate, qui a le même rôle que le NADP c’est-à-dire celui d’accepteur d’électrons.
Document 15 : L’expérience de Hill, Évolution de la quantité de dioxygène dans une suspension de chloroplastes en fonction du temps et de la lumière

Que ce soit en l’absence ou en présence de lumière, la suspension de chloroplastes isolés ne dégage pas de dioxygène mais en consomme. Il y a donc un mécanisme de respiration. La suspension de chloroplastes isolés ne dégage du dioxygène uniquement en présence de lumière qu’à partir du moment où l’on injecte dans le milieu le réactif de Hill, en l’occurrence un accepteur d’électrons. Hill en a donc déduit que l’hexacyanoferrate a accepté un électron provenant de la photolyse de l’eau. Cette déduction est confirmée par le fait qu’à l’obscurité, même en présence du réactif, il n’y a pas de libération de dioxygène mais au contraire consommation.
Ainsi en présence de lumière l’activité photosynthétique des chloroplastes dépend de la présence d’un accepteur d’électrons (un oxydant) : le NADP qui sera alors réduit.
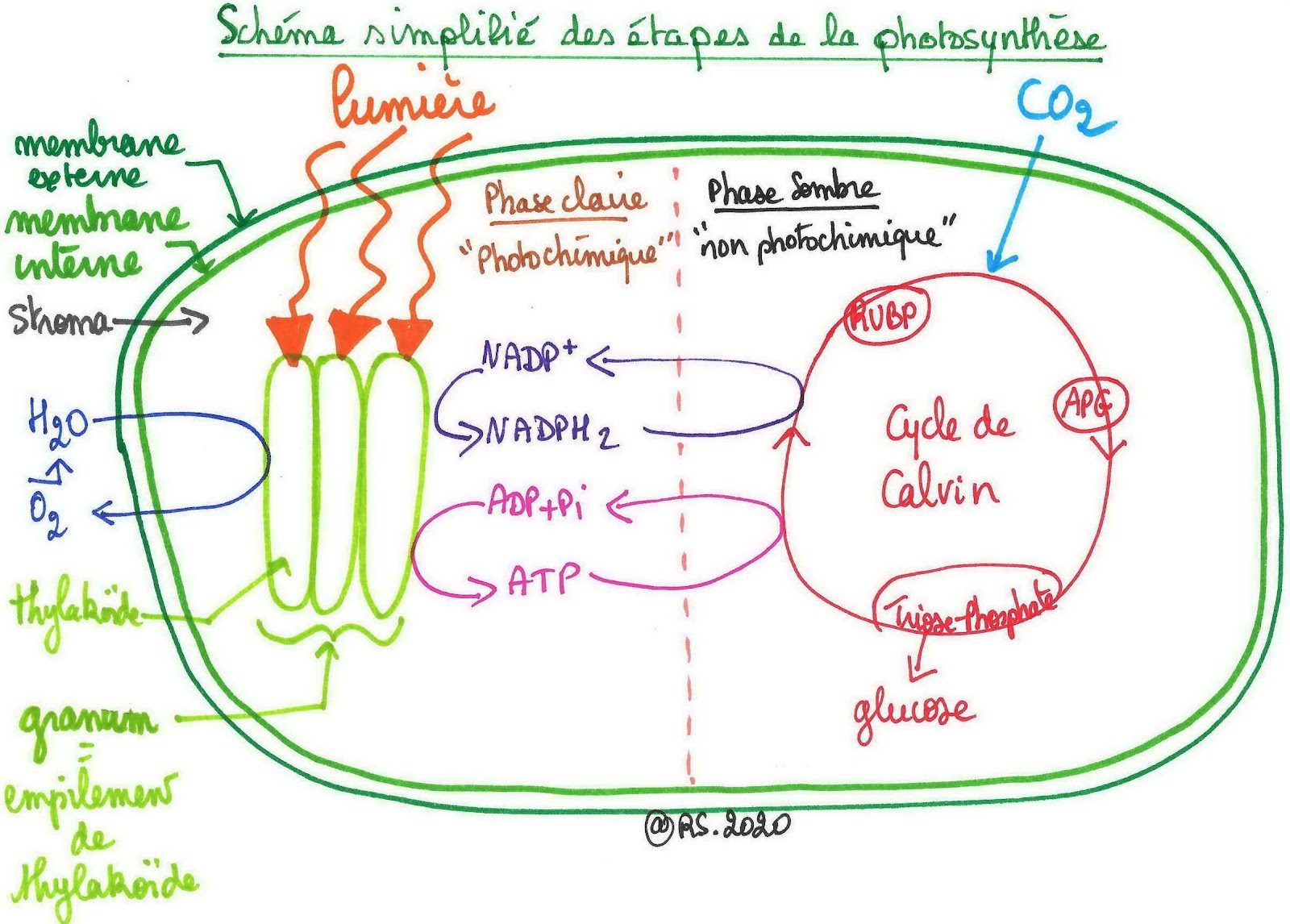
Document 16 : Schéma simplifié des étapes de la Photosynthèse
©RS.2020
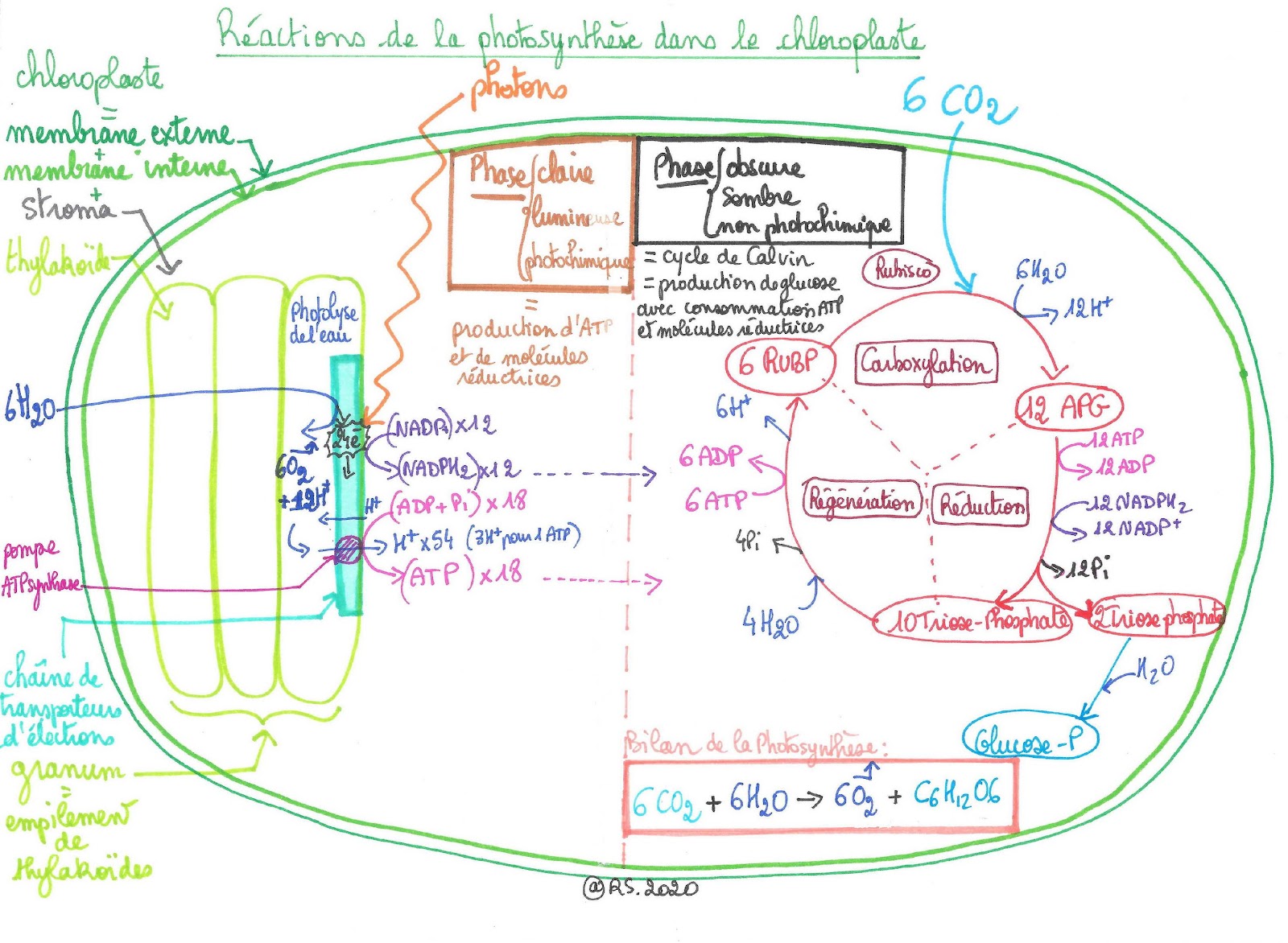
En résumé, la photosynthèse se déroule en deux étapes :
- une phase photochimique dite « claire » car nécessitant l’action de la lumière et permettant de produire l’ATP et le composé réduit NADPH2 dans les thylakoïdes du chloroplaste
- une phase chimique dite « sombre » car elle ne nécessite pas l’action de la lumière, correspondant au cycle de Calvin, se déroulant dans le stroma du chloroplaste produisant le sucre.
Il reste maintenant à comprendre qu’est-ce qui peut capter la lumière au niveau des thylakoïdes et transférer l’électron au NADP.
On sait que la photosynthèse a lieu dans les chloroplastes dont les thylakoïdes contiennent des pigments végétaux. Il est possible de séparer les pigments végétaux grâce à une chromatographie. Il faut dans un premier temps broyer des feuilles afin de récupérer une suspension des pigments puis réaliser un dépôt sur une bande de papiers Whatman que l’on dispose dans une éprouvette à chromatographie dans laquelle on aura préalablement versé un solvant. Les pigments sont très sensibles à la lumière. Il faut donc protéger le flacon de solution de pigments et la chromatographie de celle-ci le temps que le solvant migre dans le papier et les sépare. Après 20 minutes de migration on va repérer quatre tâches dont les couleurs oscillent entre le vert et le jaune.
Document 17 : protocole de la chromatographie de pigments végétaux
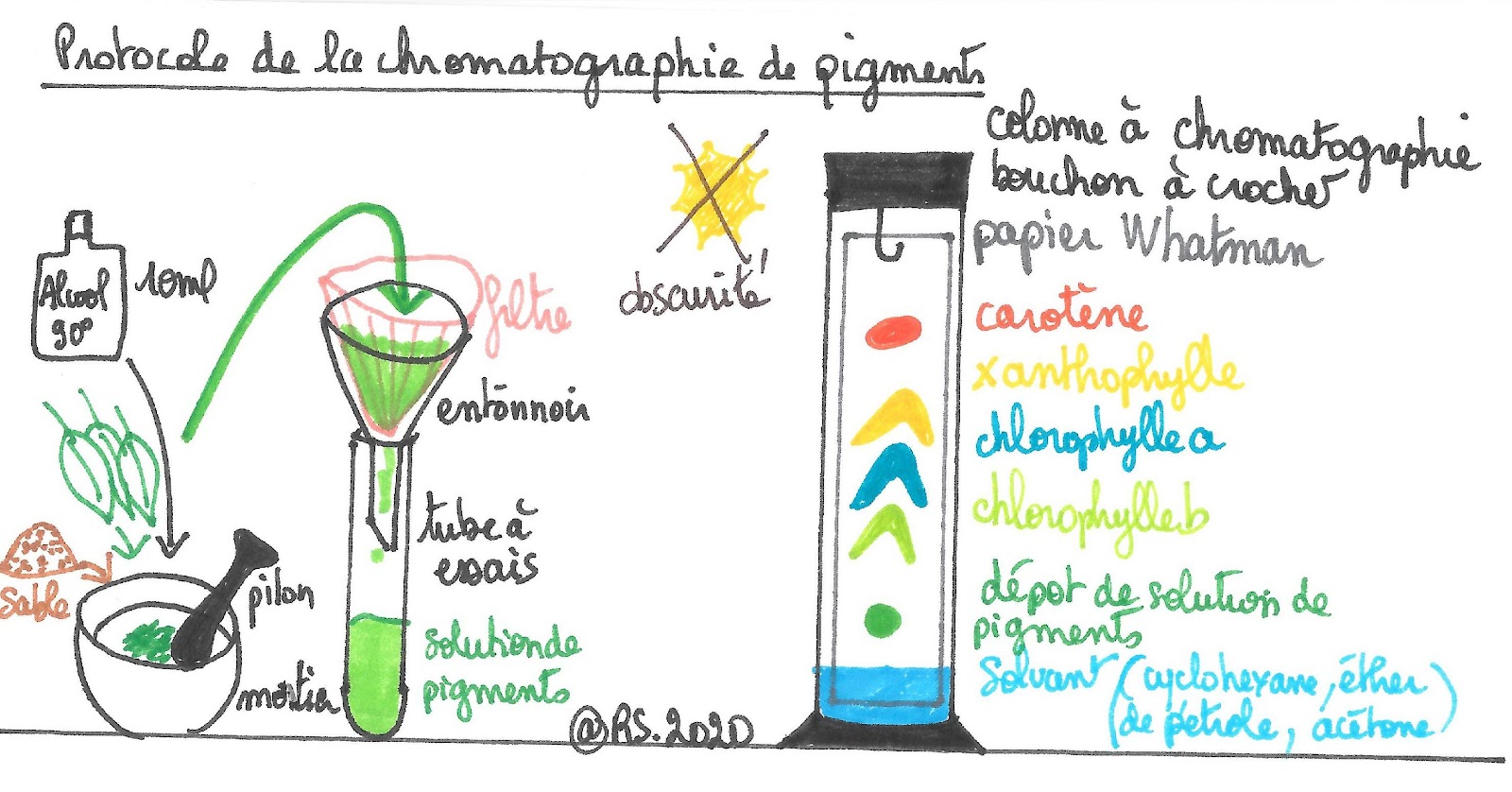
La première tâche, celle qui a le moins migré, correspond à de la chlorophylle b, de couleur vert jaune. La deuxième tâche correspond à de la chlorophylle a de couleur vert bleutée. La troisième de couleur jaune correspond au xanthophylle et la dernière jaune-orangé aux caroténoïdes.
Quatre pigments sont présents mais lesquels sont responsables de la photolyse de l’eau ?
L’analyse au spectroscope d'un faisceau de lumière traversant une solution de chlorophylle brute (solution de pigments végétaux) révèle une absorption de certaines longueurs d'ondes : les bandes noires correspondent à une absence d'ondes et donc au fait qu'elles aient été absorbées par les pigments contenus dans la cuve. Les pigments ont absorbé dans le rouge et dans le bleu puisque ces couleurs n’apparaissent plus dans le spectre de la lumière blanche.
Document 17 : Spectre d’absorption de la chlorophylle brute (mélange de pigments végétaux)
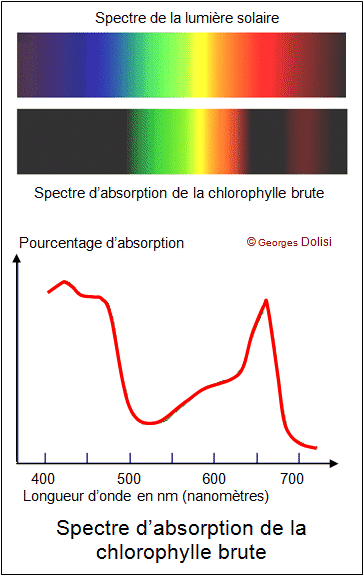
Source : Spectre chlorophylle brute.gif p ar Dolisi Georges via Wikimedia commons, CC-BY-SA-4.0, https://commons.wikimedia.org/wiki/File:Spectre_chlorophylle_brute.gif?uselang=fr
Il est possible de mesurer le spectre d’action de la photosynthèse. On mesure tout simplement chez un organisme photosynthétique, son activité photosynthétique pour différentes longueurs d’ondes qui lui sont soumises.
Document 19 : Spectre d’action photosynthétique et spectres d’absorption des différents pigments végétaux.
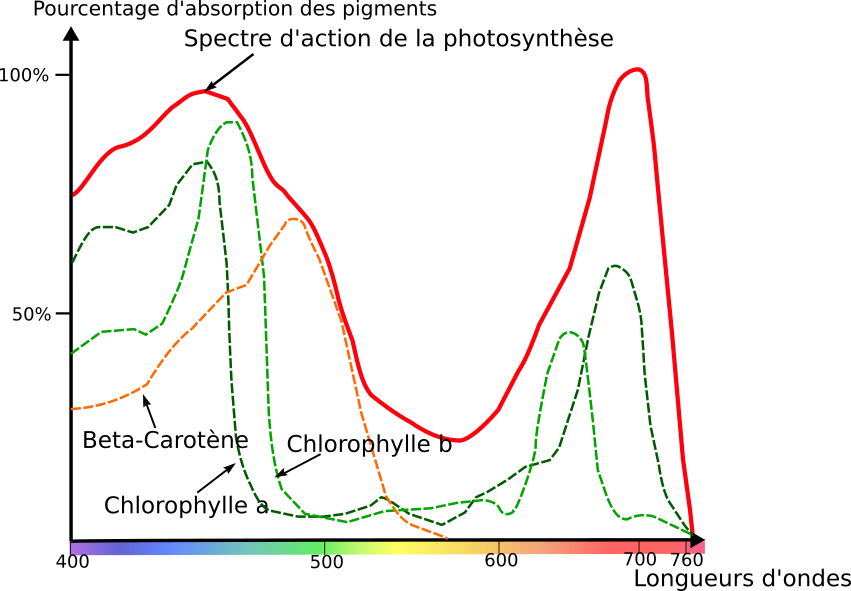
Source : Engelmannscher Bakterienversuch.svg Par Lichtabsorbtion_eines_buchenblattes.svg : Lanzi, travail dérivé: Matt ( discussion ), via Wikimédia Commons, CC-BY-SA-3.0, modifié par Sandra Rivière https://commons.wikimedia.org/wiki/File:Engelmannscher_Bakterienversuch.svg
Lorsque l’on compare le spectre d’action de la photosynthèse et le spectre d’absorption des différents pigments, on remarque que les deux spectres se superposent. L’activité photosynthétique est maximale dans les longueurs absorbées par les pigments. Ce sont donc bien les pigments qui absorbent les photons pour récupérer de l’énergie à l’origine de la photosynthèse. Les longueurs d’ondes concernées sont situées entre 450 et 500 nanomètres(le bleu) et entre 650 et 680 nanomètres (le rouge).
Ainsi on peut dire que les thylakoïdes contiennent des pigments végétaux qui vont absorber des longueurs d’onde dans le rouge dans le bleu et qui seront à l’origine de la photolyse de l’eau. Ces pigments n’absorbent pas les longueurs d’onde correspondant au vert. Ces radiations sont donc réfléchies et transmises ce qui explique la couleur verte des organes chlorophylliens. Les pigments végétaux permettent donc de convertir l’énergie lumineuse en énergie chimique (ATP).
On peut se demander comment passer d’une énergie lumineuse à une énergie chimique.
Dans les membranes des thylakoïdes sont enchâssés des photosystèmes, associations de protéines et de pigments photosynthétiques. La transformation d’énergie débute par la capture des photons par les pigments.
L’expérience d’Emerson et Arnold montre que plus un chloroplaste est éclairé, plus il photolyse de l’eau mais sans dépasser un certain seuil. Ils ont mesuré la production de dioxygène émise par une suspension de chlorelles (algues vertes unicellulaires) suite à des éclairs lumineux très brefs (inférieurs à 1 ms) envoyés en rafales d'une dizaine de secondes et d'intensité variable. La durée très brève de l'éclair laisse supposer que l'appareil photosynthétique conduisant à la libération de O2 n'est excité qu'une fois pour chaque éclair. Ils mesurent donc la production de dioxygène associée à une rafale puis ils expriment cette production par éclair (QO2/éclair) en moles O2/éclair.
Document 20 : Enregistrement de la quantité de dioxygène émise par éclair par des chlorelles contenant une quantité connue de molécules de chlorophylle en fonction de l’intensité des éclairs.

On observe que la production d’O2 augmente proportionnellement à l’intensité lumineuse de l’éclair jusqu’à atteindre un maximum qui ne change donc plus même si l’intensité lumineuse augmente. Il y a donc une valeur maximale de production d’O2 à partir d’une certaine intensité lumineuse. Comme la quantité de pigments est précise et limitée, cela signifie donc que ces pigments sont tous modifiés par la lumière : ils sont excités et vont devoir retourner un état normal avant de pouvoir à nouveau capter des photons.
Les photons sont captés par un photosystème constitué d’une « antenne collectrice de photons » elle-même constituée d’environ 2500 molécules de chlorophylles, et d’un centre réactionnel.
Document 21 : Schéma d’un photosystème
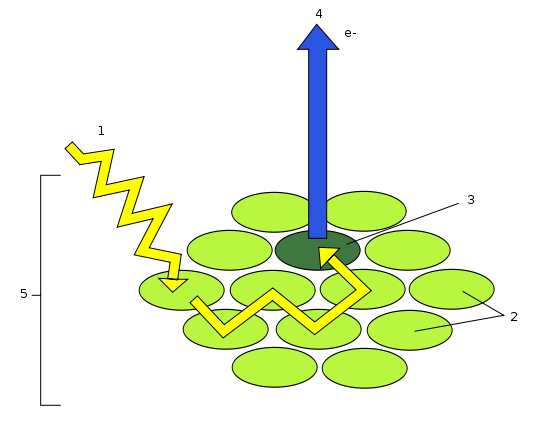
Légende : 1: Photon, 2: Molécules de pigments de l'antenne, 3: Centre réactionnel, 4: Production d'électrons vers l'accepteur primaire, 5: Photosystème
Source : Schema-photosysteme.svg par Pinpin 19:24, 24 mai 2006 (UTC)via wikimédia commons, CC-BY-SA-2.5, https://commons.wikimedia.org/wiki/File:Schema-photosysteme.svg
L'énergie des photons lumineux est captée par les différents pigments constituant l’antenne collectrice (caroténoïdes, chlorophylles a et b) puis transmise jusqu'aux molécules de chlorophylle du centre réactionnel où se réalise la réaction de séparation de charge : chaque chlorophylle excitée de ce centre va perdre un électron et passer dans un état oxydé.
Les électrons arrachés sont « excités » et parcourent toute une chaîne de transporteurs (séries de réactions d'oxydo-réduction) jusqu'à un accepteur final NADP qui est réduit en NADPH2 : les électrons riches en énergie sont donc mis en réserve dans le NADPH2. Au cours de ce transfert, il y a formation d'adénosine triphosphate (ATP) à partir d'ADP (adénosine di phosphate) et d'un Pi (phosphate inorganique) : il y a phosphorylation. L'ATP est une molécule énergétique.
Les molécules de Chlorophylle vont chercher à combler les « trous » laissés par les électrons qui ont été « arrachés » : c'est l'eau (H2O) qui va céder des électrons et des protons pour régénérer les molécules de Chlorophylle. Ce retrait scinde la molécule d'eau et permet la formation et la libération d'O2 : il y a photolyse de l’eau.
Document 22 : Chaîne de transporteurs d’électrons dans la membrane des thylakoïdes

Document 23 : Schéma bilan de la photosynthèse
{kind=link}
La photosynthèse - SVT - ENJEUX Term spé #4
Date de dernière mise à jour : 02/06/2021