Familles Multigéniques
I/ Mécanisme du crossing-over inégal
En prophase 1 de méiose, les chromosomes homologues s’apparient : ils se placent l’un à côté de l’autre et parfois leurs chromatides se chevauchent. La zone de chevauchement s’appelle un chiasma et à ce niveau les chromatides échangent des portions de matériel génétique. En temps normal l’échange est équilibré : un locus est échangé contre un même locus. On parle de crossing-over ou d'enjambement.
Quand les chromosomes homologues sont mal appariés, c'est-à-dire que le chiasma s’effectue sur des emplacements différents des chromatides homologues, l’échange ne sera pas équilibré. Une chromatide subira une perte de gène (une délétion) et l’autre subira un ajout de gène, gène qu’elle possède déjà provoquant un phénomène de duplication de celui-ci.
Document 1 : Crossing-over (enjambement)

La méiose de la cellule germinale s’effectuera donc avec une de ses paires de chromosomes qui n’est plus parfaitement homologue. La cellule-œuf résultant de la fécondation impliquant un gamète contenant le chromosome modifié présentera ainsi un génome légèrement différent de celui de son espèce. Cette cellule-œuf, au moment de sa première division, en dupliquant le chromosome simple modifié, conserve cette anomalie dans la lignée clonale qu’elle engendrera. Au fur et à mesure des divisions, les deux exemplaires du même gène vont évoluer chacun de leur côté, subissant des mutations différentes. Il va en résulter l’apparition d’un nouveau gène responsable de nouveaux caractères.
Document 2 : Comparaison des 2 chromatides homologues ayant réalisé le crossing over inégal

Source : Enjambement inégal.png, propre travail Lucquessoy, via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:Enjambement_in%C3%A9gal.png
Il arrive parfois que des échanges s’effectuent entre chromosomes non homologues sans perte ou ajout : on parle d’échange équilibré entre chromosomes non homologues ou de translocation.
Document 3 : Comparaison des 2 chromatides non homologues ayant réalisé le crossing-over inégal

Source : Déplacement 13 10.png, propre travail Lucquessoy, via Wikimédia Commons, CC-BY-SA-3.0, https://commons.wikimedia.org/wiki/File:D%C3%A9placement_13_10.png
Le gène qui a changé de chromosome se trouve donc sur un locus totalement différent. Par définition il devient donc un gène différent. Il subira lui aussi des mutations et finira par être à l’origine de nouveaux caractères. Il n'en reste pas moins que les deux exemplaires de ce gène restent proches. On parle de gènes homologues qui constituent une famille multigénique. Sont considérés comme homologues, des gènes ayant plus de 20% de similitudes dans leur séquence nucléotidique. Plus 2 gènes sont ressemblants, plus la duplication dont ils sont issus est récente.
II/ Exemples de Familles multigéniques
1) La famille des opsines
Les opsines sont des pigments des cellules des photorécepteurs de la rétine. Chaque opsine absorbe une partie spécifique du spectre lumineux : l’une absorbe dans le bleu, l’autre le rouge et la troisième dans le vert.
Le gène codant l’opsine S qui absorbe dans le bleu se situe sur le chromosome n°7 alors que les gènes codant pour les opsines M et L absorbant respectivement dans le vert et le rouge se situent sur le chromosome X.
Document 4 : Localisation des gènes des opsines

Document 5 : fichiers disponibles sur Anagène
genes-Opsines.edi : comparaison des 3 gènes d’opsines chez l’Homme
OpsPrimates.edi : comparaison des séquences protéiques d’opsine S entre primates
ops_callithrix_cebus_saimiri_pro.edi : comparaison séquences protéiques des opsines S et M/L des singes nouveau monde
La comparaison sous Anagène de leur séquence codante permet de tracer un tableau appelé matrice des identités. Dans ce tableau, la valeur inscrite dans une case donne le pourcentage de similitude entre le gène inscrit en titre de ligne et le gène inscrit en titre de colonne.
Pour obtenir ce pourcentage d’identité il suffit de lancer une comparaison avec discontinuité entre un gène mis en référence et un autre gène, puis dans la fenêtre de comparaison, sélectionner le gène étudié et dans la barre des tâches appuyer sur le bouton « i » pour information. Une fenêtre s’ouvre et l’identité vous est indiquée.
Document 22 : Recherche d’identité entre le gène de l’opsine bleue et le gène de l’opsine verte mise en référence sous Anagène

Il suffit donc tour à tour de placer chaque gène en référence et de comparer les séquences pour obtenir les pourcentages d’identité entre chacun des gènes
Document 6 : Matrice des identités pour les gènes des opsines
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
Les grandes similitudes entre les séquences des pigments M et L, tant au niveau protéique que nucléique, indiquent une parenté importante entre ces molécules. Les deux gènes M et L sont donc issus d'un même gène ancestral : il y a eu duplication suivie de mutations, certaines au moins de ces mutations étant à l'origine des différences de spectre d'absorption entre les pigments codés par ces gènes, ce qui contribue à la vision des couleurs. L’opsine S a plus de 50 % de similitudes avec les gènes M et L, ce qui permet d'envisager aussi une origine commune pour tous ces gènes. Au cours de l'histoire des gènes des opsines, qui forment donc une famille multigénique, trois duplications sont donc à envisager, suivies de mutations.
Document 7 : Arbre d’évolution des opsines

Pour comprendre l’origine des trois opsines de l’Homme, retraçons leur histoire en comparant leurs séquences protéiques sous anagène avec les séquences protéiques des opsines des primates.
Rappelons qu’il existe 2 groupes de primates : ceux de l’ancien monde (Europe, Asie) et ceux du nouveau monde (Amérique). Les deux lignées se sont séparées il y a 34 millions d’années. Les singes du nouveau monde appartiennent au groupe des Platyrrhini qui signifie « nez large» et qui ont des narines fendues. Les singes de l’ancien monde appartiennent au groupe des Catarrhini qui signifie « nez bas » et qui ont des narines fermées et rapprochées et dont l’Homme fait partie.
L’étude sous Anagène des séquences en acides aminés de l’opsine S montre qu’il y a de très grandes similitudes entre les opsines des singes de l’ancien monde (Bonobo, chimpanzé, gorille, macaque). Le singe le plus éloigné de l’Homme est le macaque avec 96 % d’identité. L'ancêtre commun à l'Homme et au Macaque
possédait donc déjà ce système génétique.
Document 8 : Comparaison de l’opsine S de l’Homme et du Macaque sous Anagène
possédait donc déjà ce système génétique.
Document 25 : Comparaison de l’opsine S de l’Homme et du Macaque sous Anagène

Les singes du nouveau monde (alouate, saimiri, cebus) ont un système génétique différent. S’ils possèdent le gène de opsine S sur le chromosome 7, sur leur chromosome X, il n'existe qu'un seul gène codant pour un pigment appelé opsine M/L ayant un spectre d'absorption présentant un maximum à plus de 500 nm (au lieu de deux gènes M et L chez le Macaque et l'Homme).
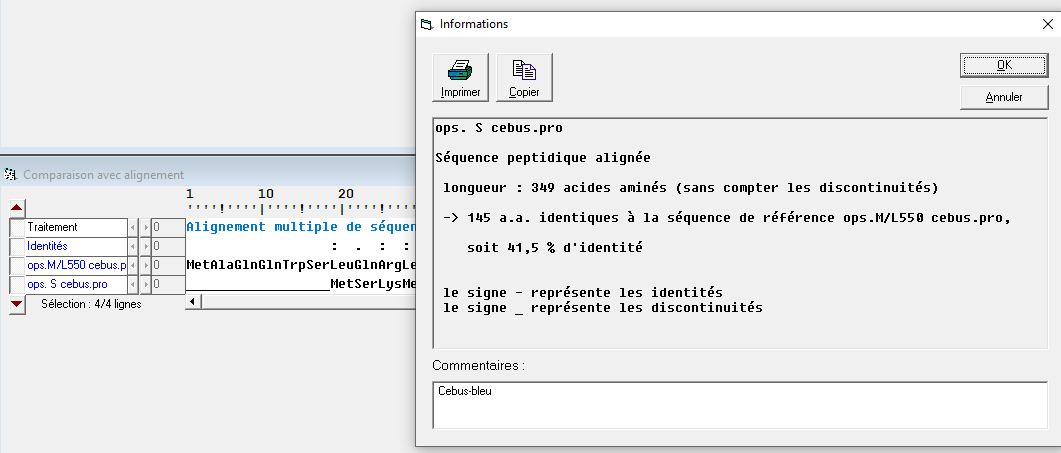
La comparaison des séquences protéiques de l’opsine S et de l’opsine M/L chez Cebus montre 41.5% d’identité. Ce sont donc deux gènes homologues issus de la duplication d’un gène ancestral.
Document 9 : Comparaison de l’opsine S et de l’opsines M/L des singes du nouveau monde sous Anagène
L’histoire de la famille commence à se dessiner. Chez l’ancêtre de tous les primates, il n’existait qu’un seul gène responsable de la fabrication d’une opsine sur un chromosome. Suite à un crossing-over entre deux chromosomes non homologues, il s’effectue une duplication du gène. Deux chromosomes portent alors un gène d’opsine : le n°7 et le X.
L’opsine S du chromosome n°7 de l’Homme présente 92.5 % d’identité avec celle de Cebus singe du nouveau monde. Il y a donc eu chez les singes de l’ancien monde des mutations à l’origine de l’opsine S de ceux-ci.
Document 10 : Comparaison de l’opsine S de l’Homme et de l’opsine de Cebus sous Anagène

On a vu que la comparaison des opsines M et L de l’Homme donnait 97.7% d’identité. On peut donc affirmer que chez les singes de l’ancien monde, il y a eu un crossing-over inégal entre les deux chromosomes X amenant à une duplication du gène. Chaque exemplaire a alors subit des mutations ce qui a abouti aux 2 gènes M et L sur X. Cette duplication à l'origine des gènes M et L a eu lieu après la séparation des singes du nouveau monde /singes de l'ancien monde, soit entre -40 et -23 Ma.
À partir de toutes ces informations, on peut retracer les mécanismes génétiques à l’origine des différents gènes d’opsines.
Document 11 : Mécanismes génétiques à l’origine de la famille multigénique des opsines

2) La famille des globines
Les globines sont des protéines impliquées dans le transport ou le stockage du dioxygène. Il en existe plusieurs sortes. Elles sont l’unité de bas de l’Hémoglobine. Chaque molécule d'hémoglobine est un tétramère formé par l'association de quatre chaînes polypeptidiques identiques deux à deux. Chaque chaîne adopte une conformation spatiale lui donnant une forme globuleuse et ménageant une "poche" superficielle dans laquelle se trouve logé un hème contenant un ion Fe2+ capable de fixer les molécules de dioxygène.
Document 12 : Molécule d’hémoglobine observée sous Rastop constituée de 2 globines alpha en rouge et 2 globines bêta en bleu.

Source : Hemoglobin-3D-ribbons.png, par Benjah-bmm27 via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Haemoglobin-3D-ribbons.png
Document 13 : Évolution des proportions des différentes globines dans le sang avant et après la naissance.
Source : Globines humaines à la naissance.svg, par Léonid 2 via Wikimédia Commons, domaine publique, https://commons.wikimedia.org/wiki/File:Globines_humaines_%C3%A0_la_naissance.svg
Il existe plusieurs globines ce qui implique qu’il existe plusieurs gènes. Ces gènes sont situés sur deux chromosomes différents : le chromosome 11 et le chromosome 16.
Document 14 : Localisation des gènes des globines

Les séquences nucléotidiques ou les séquences peptidiques des globines humaines présentent un degré de similitude qui ne peut être dû au hasard. Les globines sont donc des protéines homologues ainsi que les gènes qui les codent. Tous ces gènes dérivent d'un même gène ancestral. Cela signifie qu’il y a eu un évènement de crossing-over entre 2 chromatides non homologues et que cela a amené à la duplication du gène ancestral.
Document 15 : Matrice des identités pour les gènes des globines
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
Les deux globines les plus proches sont les globines delta et bêta. Elles dérivent donc d'un même gène ancestral.
Ensuite on remarque que le gène le plus proche est gamma. Il est un peu plus éloigné de delta que de bêta ce qui signifie que le groupe "bêta et delta" a un ancêtre commun avec gamma mais que c'est delta qui a le plus muté.
Enfin alpha est celui qui possède le pourcentage d'identité le plus faible mais quand même supérieur à 20 %, c'est donc un gène homologue issu d'une duplication à partir d'un gène ancestral commun à tous.
Document 16 : Arbre d’évolution des globines

Les gènes des globines forment donc une famille multigénique dont on peut reconstituer les mécanismes évolutifs grâces aux pourcentages d’identité.
Document 17 : Mécanismes génétiques à l’origine de la famille multigénique des globines

Familles multigéniques-SVT - LA VIE Term spé #9 - Mathrix
Date de dernière mise à jour : 26/05/2021